
- Орангутаны научились использовать орудия с выгодой для себя [2019-03-22]
- Мозг собак отличил настоящие слова от тарабарщины [2019-03-22]
- Какаду Гоффина выклевали палочки из картонки и использовали их в быту [2019-03-22]
- Орангутанов уличили в создании крючков [2019-03-22]
- Любовь самок гуппи к ярким самцам объяснили генами и освещением [2019-03-22]
- Орангутаны рассказывают друг другу о прошлом [2019-03-22]
- Блеск и нищета этологии [2018-05-03]
- Почему хозяева похожи на своих собак? [2018-03-15]
- Интервью с Анатолием Протопоповым [2019-02-13]
- Цирки: развлечение vs мучение [2018-12-15]

Вывод из запоя на дому в москве
Вывод из запоя на дому с выездом в москве вывод из запоя на дому в москве.
germes-narkologiya.ru
Э.О. Уилсон
Человек: от социобиологии к социологии
(перевод: А. Протопопов, М. Потапов, А. Вязовский, С. Разумная)
Давайте рассмотрим человека в вольном духе естествознания, как будто бы мы – зоологи с другой планеты, составляющие каталог социальных видов на Земле. При таком отстраненном взгляде гуманитарные и общественные науки “сжимаются” до специализированных отделов биологии; история, биографии и беллетристика выглядят просто как протоколы этологии человека; а антропология вместе с социологией составляют частную социобиологию конкретного вида приматов.
В экологическом отношении Homo sapiens – совершенно особый вид. Он имеет самую географически обширную область распространения и сохраняет самую высокую локальную плотность населения по сравнению со всеми прочими приматами. Мудрый инопланетный эколог не удивился бы, обнаружив, что в роде Homo существует только один вид, поскольку современный человек занял все мыслимые экологические ниши гоминид. В прошлом же сосуществовали два или более видов гоминид, и в дополнение к ранним представителям Homo в Африке жили человекоподобные обезьяны – австралопитеки (Australopithecus). Однако до позднего плейстоцена дожила только одна эволюционная ветвь, которая и причастна к появлению наиболее совершенных социальных черт человека.
Современный человек уникален в анатомическом плане. Другие приматы, включая гориллу и шимпанзе, лишь изредка ходят на задних ногах, даже отдаленно не достигая его вертикального положения тела с полным переходом к хождению на двух конечностях. В связи с прямохождением, строение человеческого скелета подверглось существенным изменениям. Так, позвоночный столб приобрел изгибы для более равномерного распределения веса тела вдоль его длины; грудь уплощилась, чтобы сместить центр тяжести назад, к позвоночнику. Таз расширился для крепления мощных шагательных мышц бедра и превратился в чашу для поддерживания внутренностей; хвост исчез, а его позвонки (теперь называемые копчиком) изогнулись вовнутрь и сформировали часть дна тазовой чаши. Затылочные мыщелки переместились далеко в нижнюю часть черепа, в результате чего вес головы уравновешен на них; лицевая часть черепа сократилась, способствуя такому перераспределению нагрузки. Большой палец руки увеличился, что придало ей силу. Нога удлинилась; ступня чрезвычайно сузилась и также удлинилась для облегчения ходьбы. Произошли и другие изменения, в частности, на большей части тела исчезли волосы. Почему современный человек превратился в “голую обезьяну” до сих пор неизвестно. Одно из вероятных объяснений этого факта состоит в том, что нагота способствовала охлаждению тела во время напряженного преследования добычи в жарких условиях Африканских равнин. Дело в том, что человек использует исключительно потоотделение для снижения температуры тела, а человеческое тело содержит от двух до пяти миллионов потовых желез – гораздо больше, чем у любого другого вида приматов.
Репродуктивная физиология и поведение Homo sapiens также претерпели удивительную эволюцию. В частности, эстральный цикл женщины изменился в двух аспектах, затрагивающих половое и социальное поведение. Усилилась менструация, и если самки других приматов испытывают лишь небольшое кровотечение, то только у женщин происходит интенсивное обрушение стенки “разочаровавшейся матки” с последующим интенсивным кровотечением. Эструс, или период “течки”, у женщин заместился фактически непрерывной половой активностью. Совокупление инициируется не в ответ на обычные для приматов признаки эструса, такие как изменение цвета кожи вокруг женских половых органов и выделение феромонов, а в результате длительной предшествующей игры, приводящей партнеров в обоюдное возбуждение. Кроме того, зафиксировались признаки, вызывающие физическое влечение, такие как волосы на лобке у обоих полов и выпуклые груди и ягодицы у женщин. Сглаженный половой цикл и непрерывная женская привлекательность цементируют узы брака, которые составляют основу социальной жизни человека.
Глядя со стороны, проницательный марсианский зоолог расценил бы шаровидную голову как важнейший ключ к человеческой биологии. Головной мозг Homo чрезвычайно увеличился в течение относительно короткого промежутка эволюционного времени (см. Рис. 26-1). Три миллиона лет назад объем черепа взрослого австралопитека составлял 400–500 куб. см, что сопоставимо с таковым шимпанзе и гориллы. Два миллиона лет спустя его предполагаемый потомок Homo erectus имел мозг с объемом уже около 1000 куб. см. В течение следующего миллиона лет произошло его увеличение до 1400–1700 куб. см у неандертальца и 900–2000 куб. см у современного Homo sapiens. Рост интеллекта, который сопровождал это увеличение, был настолько большим, что его невозможно оценить каким-либо рациональным способом. Можно сравнить между собой людей по нескольким основным компонентам интеллекта и творческого потенциала, но шкала для объективного сравнения человека с шимпанзе и другими ныне живущими приматами пока не придумана.
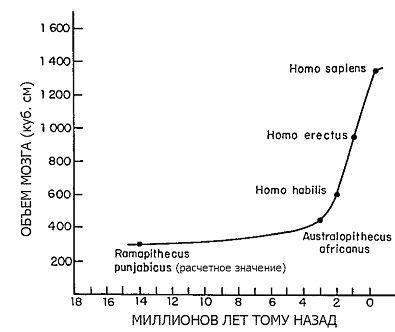 |
| Рисунок 26-1. Увеличение размера мозга в ходе эволюции человека (Воспроизведено из Pilbeam, 1972). |
Мы совершили столь стремительный эволюционный скачок в развитии интеллекта, что он пока не поддается нашему анализу. Гипертрофия интеллекта привела к искажению почти до неузнаваемости даже самых основополагающих социальных характеристик, типичных для приматов. Разные виды низших и высших обезьян Старого Света обладают высокой пластичностью социальной организации, человек же распространил эту тенденцию в многоликую расово-этническую область. Для регулирования агрессивных и половых взаимодействий обезьяны используют поведенческое шкалирование (приведение формы или интенсивности реакции в соответствие с величиной стимула – прим. перев. М.П.), у человека же сами эти шкалы стали многомерными, модифицируемыми в зависимости от культуры и практически бесконечно тонко настраиваемыми. Тесная взаимосвязь особей и практика реципрокного альтруизма у других приматов находятся в зачаточном состоянии, человек же расширил их применение на огромные взаимосвязанные общности, где индивидуумы сознательно меняют свои роли чуть ли не ежечасно, словно маски.
В задачу сравнительной социобиологии входит максимально детально проследить эти и другие человеческие качества в исторической аспекте. Помимо привнесения перспективы и, возможно, некоторого ощущения философской свободы, это занятие поможет определить формы поведения и правила, с помощью которых человеческие индивидуумы увеличивают дарвиновскую приспособленность путем манипулирования сообществом. Иначе говоря, мы хотим проанализировать человеческую биограмму (Count, 1958; Tiger and Fox, 1971). Один из ключевых вопросов, никогда не оставляющих антропологов и биологов, занимающихся истинной теорией, состоит в том, до какой степени биограмма представляет адаптацию к современной культурной жизни и до какой степени это филогенетическое наследство. Наши цивилизации были построены в какой-то мере вокруг биограммы. Как она влияла на них? С другой стороны, насколько биограмма гибка и по каким параметрам в особенности? Опыт изучения других животных свидетельствует о том, что гипертрофия органов затрудняет реконструкцию их филогении. Это определяет основную проблему эволюционного анализа человеческого поведения. В оставшейся части главы мы обсудим человеческие качества в той мере, в какой они являются общими чертами вида. Затем будет сделан обзор текущего состояния знаний об эволюции биограммы, и, наконец, рассмотрены некоторые выводы – для планирования будущих обществ.
Пластичность социальной организации
Первая и наиболее легко проверяемая диагностическая характеристика – статистическая по природе. Параметры социальной организации, включая размер группы, свойства иерархий и скорость обмена генами, в гораздо большей степени варьируют между популяциями человека, чем любых других видов приматов, превосходя даже их межвидовую изменчивость. Некоторого увеличения пластичности и следовало бы ожидать, экстраполируя тенденцию к вариабельности, очевидную уже у павианов, шимпанзе и других обезьян Старого Света; удивительна, однако, та высокая степень, до которой она доведена.
Почему же человеческие сообщества столь гибки? Отчасти причина в том, что сами их члены очень изменчивы по своему поведению и способностям. Индивидуумы весьма отличаются друг от друга даже в самых просто устроенных обществах. В маленьком бушменском племени Кунг находятся признаваемые “лучшими людьми” лидеры и выдающиеся специалисты среди охотников и целителей. Более того, некоторые из них, имеющие склонность к обмену товарами, оказываются исключительно способными предпринимателями и потихоньку скапливают некоторое богатство. Люди Кунг, также как люди современных индустриальных обществ, обычно самоутверждаются где-нибудь годам к тридцати пяти, в противном же случае смиряются с меньшим жизненным статусом. Некоторые никогда не стремятся к самоутверждению и живут себе в ветхих хижинах, не испытывая гордости ни за себя, ни за свой труд (Pfeiffer, 1969). Способность переходить к таким ролям, соответствующим личностным качествам, сама по себе может быть адаптивной. Человеческие общества основаны на сообразительности, а каждый их член стоит перед лицом целого комплекса социальных проблем, бросающих вызов всей его находчивости. Эта исходная индивидуальная изменчивость усиливается на уровне групп благодаря исключительно развитым в человеческих сообществах качествам, таким как длинный и напряженный период социализации, отсутствие жесткости коммуникационных сетей, разнообразие индивидуальных связей, способность (особенно в культурах с развитой грамотой) преодолевать большие расстояния и целые периоды истории для передачи информации и, вследствие всего перечисленного, способность притворяться, подтасовывать, преследуя корысть, и эксплуатировать других. Каждый параметр легко изменяем, и каждый имеет заметное влияние на окончательную социальную структуру, в результате чего мы и наблюдаем всю изменчивость сообществ.
Из этого следует, что достойна рассмотрения гипотеза о том, что гены, обеспечивающие гибкость социального поведения, прошли мощный отбор на индивидуальном уровне. Заметим, однако, что изменчивость социальной организации может не быть необходимым следствием этого процесса. Чтобы сгенерировать фактически наблюдаемое число вариантов, необходимо существование множественных адаптивных пиков. Другими словами, для продолжительного благополучного сосуществования различных форм сообществ в пределах одного вида необходимо, чтобы они давали сходные шансы на выживание для множества их членов. Результатом этого был бы статистический ансамбль типов сообществ, если не равновесный, то, по крайней мере, не сдвигающийся слишком быстро к одному или другому конкретному способу.
Альтернатива, найденная некоторыми общественными насекомыми, – это гибкость индивидуального поведения при одновременном развитии кастовости, которые, не смотря ни на что, приближает статистическое распределение категорий особей совокупной колонии к однотипности. У медоносных пчел и муравьев родов Formica и Pogonomyrmex, “личностные” различия – даже в пределах отдельных каст – сильно выражены. Некоторые особи, относимые энтомологами к элите, необычайно активны, делают больше, чем им полагается за свою жизнь, да еще и других побуждают работать, оказывая им вспомоществование. Другие члены колонии в той же мере бездеятельны. Хотя на вид они здоровы и долго живут, их индивидуальная производительность составляет лишь малую долю таковой элитных особей. Наблюдается также специализация: определенные особи остаются с выводком в качестве нянек значительно дольше среднего, в то время как другие концентрируют свои усилия на гнездостроении и фуражировке. И все же, каким-то образом целостная структура поведения в колонии сводится к средней, типичной для данного вида. Если сравнить одну колонию, состоящую из сотен и тысяч особей, с другой того же вида, оказывается, что структуры активности у них статистически сходны. Известно, что отчасти такое соответствие обеспечивается отрицательными обратными связями. Когда усиливается потребность в заботе о выводке или ремонте гнезда, рабочие особи изменяют свою активность таким образом, чтобы эту потребность компенсировать, а затем возвращаются к прежним делам. Показано экспериментально, что разрушение обратных связей и, как следствие, отклонение колонии от статистической нормы может быть гибельным. Потому и неудивительно, что петли обратных связей столь точны и прочны (Wilson, 1971a).
Механизмы, управляющее человеческими сообществами, далеко не так сильны, а эффекты отклонений не настолько опасны. Антропологическая литература изобилует примерами обществ, отягощенных явной неэффективностью и патологическими пороками, которые, однако, выдерживают испытание временем. Рабовладельческое общество Ямайки, ярко описанное Орландо Паттерсоном (Patterson, 1967), с точки зрения нравственных норм цивилизованной жизни, было бесспорно нездоровым. “Что отличает его, так это поразительное пренебрежение и нарушение почти каждой из основных предпосылок нормального человеческого существования. Это было общество, в котором священнослужители были самыми законченными распутниками на земле; в котором институт брака официально порицался как среди рабовладельцев, так и среди рабов; в котором семья была немыслима для подавляющего большинства населения, а промискуитет был нормой; в котором образование считалось пустой тратой времени, а от учителей бежали, как от чумы; в котором правовая система была преднамеренно превращена в пародию на то, что можно назвать правосудием; в котором все формы изысканности, искусств, народных обычаев или отсутствовали или находились в состоянии полного разложения. Только малая доля белых людей, монополизировавших почти всю плодородную землю острова, извлекала выгоду из этой системы. И они, обеспечив свое будущее, покинули эту землю, из которой они выжали все, сделав непригодной для проживания, ради того, чтобы обрести комфорт и благоденствие в отчизне”. И все же этот гоббесианский мир (“война всех против всех” – прим. перев. М.П.) продолжался почти два столетия! Люди умножались, в то время как экономика процветала.
Как может сохраняться такая изменчивость социальной структуры? Объяснением может служить отсутствие конкуренции со стороны других видов, приводящее к тому, что биологи называют экологическим высвобождением. В течение прошедших десяти или более тысяч лет человек как целое был настолько успешен в доминировании над средой, что почти любой вид культуры мог преуспевать, если имел даже самую скромную степень внутренней связности и не отключал полностью воспроизводство. Никакой вид муравьев или термитов не наслаждается такой свободой. Самая небольшая степень неэффективности при построении гнезд, в прокладывании запаховых троп или в проведении брачных полетов могла бы привести к быстрой гибели вида из-за хищничества и конкуренции со стороны других общественных насекомых. В несколько меньшей степени, но это справедливо и для социальных видов хищников и приматов. Короче говоря, любой вид животных плотно “упакован” в экосистеме и имеет очень небольшую возможность для экспериментов или игр. Человек пока избежал ограничений, связанных с межвидовой конкуренцией. Хотя культуры сменяют друг друга, процесс этот намного менее эффективен в деле сокращения изменчивости, чем межвидовая конкуренция.
Общепринято считать, что фактически вся культурная изменчивость имеет скорее фенотипический, чем генетический источник. Это представление подтверждается той легкостью, с какой некоторые аспекты культуры изменяются в течение одной смены поколений – слишком быстро, чтобы иметь эволюционную природу. Решительное изменение в ирландском обществе за первые два года “картофельной напасти” (1846–1848 гг.) – типичный случай (уничтожение урожая картофеля – основного продукта питания ирландцев картофельной тлей привело к падению народонаселения страны с 9 до 4 млн. человек вследствие смертности от голода и эмиграции – прим. перев. С.Р.). Другой – изменение структуры власти в Японии за время американской оккупации после второй мировой войны (под влиянием оккупации была принята конституция, образцом для которой послужила парламентская демократия англосаксонского типа, хотя во многом страна сохранила традиционные ценности, нормы группового сотрудничества и принципы патернализма – прим. перев. С.Р.). Такие примеры можно множить бесконечно, они – сущность истории.
В своем крайне ортодоксальном представлении энвайронментализм заходит еще дальше, утверждая, что в действительности в передаче культуры не содержится генетической составляющей. Другими словами, способность к усвоению культуры передается единственно человеческим генотипом. Добржанский (Dobzhansky, 1963) сформулировал эту гипотезу следующим образом: “Культура не наследуется через гены, она приобретается через обучение от других людей … В некотором смысле, человеческие гены уступили первенство в человеческой эволюции абсолютно новому, небиологическому или надорганическому средству – культуре. Однако не должно забывать, что это средство полностью зависит от человеческого генотипа”. Хотя гены отдали большую часть владычества, они сохраняют определенное влияние, по крайней мере, на поведенческие качества, лежащие в основе изменчивости между культурами. Умеренно высокая наследуемость зарегистрирована для показателей интро/экстровертности, личного темпа жизни, психомоторной и спортивной активности, невротичности, доминирования, склонности к депрессиям, возраста первого полового контакта, сроков основного когнитивного развития и склонности к некоторым формам психических расстройств типа шизофрении (Parsons, 1967; Lemer, 1968; Martin et al., 1977; R. Wilson, 1979). Даже малая часть этой дисперсии, перенесенная на межпопуляционные различия, может предрасположить сообщества к возникновению различий в культуре. По крайней мере, мы должны попробовать измерить эту часть. Безосновательно указывать на отсутствие какой-либо поведенческой особенности в одном или нескольких обществах как на решающее доказательство того, что данная особенность индуцируется окружающей средой и никак не связана с генетической предрасположенностью в человеке. Истиной может оказаться совершенно обратное.
Действительно, Фридман (Freedman, 1974, 1979) и его коллеги продемонстрировали отчетливые расовые различия в характере движений, осанке, мышечном тонусе и эмоциональной реакции новорожденных младенцев, которые нельзя приемлемо объяснить обучением или даже выработкой условного рефлекса во чреве. Среди американских новорожденных этнические китайцы, например, более психически устойчивы, меньше тревожатся от шума и движения, лучше приспосабливаются к новым стимулам и дискомфорту и быстрее успокаиваются, чем младенцы европейской расы. Точнее говоря, случайная выборка младенцев, чьи предки происходят из определенных мест Китая, отличается по указанным поведенческим особенностям от сопоставимой выборки европейского происхождения.
Несомненно, нам нужна такая дисциплина, как антропологическая генетика. Прежде, чем она появится, охарактеризовать человеческую биограмму можно с помощью двух косвенных методов. Во-первых, на основе простейших норм человеческого поведения можно создать модели. В той мере, в которой их можно протестировать, эти нормы характеризуют биограмму аналогично тому, как этограммы, составленные зоологами, идентифицируют “типичные” поведенческие репертуары различных видов животных. Значит, на законных основаниях человеческие нормы поведения можно сравнить с этограммами других видов приматов. Изменчивость норм среди человеческих культур, хоть и небольшая, могла бы дать ключи к пониманию основных генетических различий, особенно, когда она коррелирует с изменчивостью заведомо наследуемых поведенческих особенностей.
Социологи фактически начали брать на вооружение этот первый подход, хотя и в другом контексте, чем тот, что предложен здесь. Авраам Маслоу (Maslow, 1954, 1972) постулировал, что люди отвечают на иерархию потребностей таким образом, что более низкие уровни удовлетворяются прежде, чем большое внимание будет уделено более высоким. Самые основные потребности – утоление голода и сон. Когда они удовлетворены, первое место занимает безопасность, затем необходимость принадлежать к группе и получать сексуальное удовлетворение. За этим следует чувство собственного достоинства, и, наконец, самореализация и творчество. Идеальное общество в мечтах Маслоу это то, которое “стимулирует наиболее полное развитие человеческого потенциала, полной степени человечности”. Когда реализации биограммы ничто не мешает, центр ее тяжести должен успокоиться на более высоких уровнях. Второй социолог, Джордж Каспар Хоманс (Homans, 1961), принял скиннеровский подход в попытке расчленить человеческое поведение до основных процессов ассоциативного обучения. Правила, которые он постулировал, таковы:
1. Если в прошлом, в случае наступления конкретной побудительной ситуации, деятельность человека была вознаграждена, тогда чем более побудительная ситуация в настоящем подобна таковой в прошлом, тем более вероятно, что человек в настоящий момент совершит такую же или подобную деятельность.
2. Чем чаще в данный период времени определенная деятельность человека вознаграждает поведение другого, тем чаще этот другой будет демонстрировать такое же поведение.
3. Чем более ценна для человека единица направленной на него деятельности со стороны другого, тем чаще он ведет себя так, чтобы быть вознагражденным этой деятельностью другого.
4. Чем чаще человек в недавнем прошлом получал вознаграждающую деятельность от другого, тем менее ценной становится для него каждая дальнейшая единица этой деятельности.
Маслоу, этолог и мечтатель, кажется полной противоположностью Хомансу, бихевиористу и редукционисту. И все же, их подходы совместимы. Правила Хоманса можно рассматривать как содержащие некоторые механизмы для изображения человеческой биограммы. Его рабочее слово – вознаграждение, которое является фактически набором всех взаимодействий, определяемых эмоциональными центрами мозга как приятные и желательные. Согласно эволюционной теории, желательность измеряется в единицах генетической приспособленности; соответственно были запрограммированы и эмоциональные центры. В таком случае, иерархия Маслоу – просто порядок очередности целей, на достижение которых ориентированы правила.
Второй косвенный подход к антропологической генетике – филогенетический анализ. При сравнении человека с другими видами приматов становится возможным идентифицировать их лежащие под поверхностью основные особенности, помогающие разобраться в конфигурации более высокого социального поведения человека. Этот подход применен в серии популярных книг, написанных хорошим слогом и с большой энергией Конрадом Лоренцом (“Агрессия”; Konrad Lorenz, On Aggression), Робертом Ардри (Robert Ardrey, The Social Contract), Десмондом Моррисом (“Голая обезьяна”; Desmond Morris, The Naked Ape), а также Лайонелом Тайгером и Робином Фоксом (Lionel Tiger and Robin Fox, The Imperial Animal). Их усилия были благотворны, ибо привлекли внимание общества к статусу человека как биологического вида, адаптированного к специфической среде. Широкий интерес, который вызвали их работы, разорвал удушающую хватку крайних бихевиористов, представление которых о человеке фактически как об эквипотенциальной реагирующей машине не было ни правильным, ни эвристическим. Однако особое обращение указанных авторов к проблеме было чревато малой эффективностью и введением в заблуждение. Они выбирали ту или иную гипотезу на основе обзора слишком малого представительства животных видов, а затем упорно настаивали на своих объяснениях.
Правильный же подход с использованием сравнительной этологии должен основывать строгую филогению близкородственных видов на многих их биологических особенностях. В таком случае, социальное поведение рассматривается как зависимая переменная и с учетом этого прослеживается его эволюция. Когда этого нельзя сделать с уверенностью (и нельзя сделать для человека), следующей наиболее оправданной процедурой должно стать установление самого низкого таксономического уровня, на котором каждый признак демонстрирует наиболее значимую межтаксонную изменчивость. Признаки, которые изменяются от вида к виду или от рода к роду, являются наиболее лабильными. Мы не можем безошибочно экстраполировать их с обезьян Старого Света на человека. У приматов такими лабильными качествами являются размер группы, связность группы, открытость группы к другим, участие самца в родительской заботе, структура внимания, а также интенсивность и форма защиты территории. Признаки считаются консервативными, если они сохраняют постоянство на уровне таксономического семейства или в пределах всего отряда Приматов, и тогда они наиболее вероятно сохраняются в относительно неизменной форме в эволюции Homo. Эти консервативные особенности включают системы агрессивного доминирования, где мужчины обычно доминируют над женщинами; шкалирование интенсивности реакций, особенно – агрессивных взаимодействий; интенсивная и длительная материнская забота с выраженной степенью социализации молодых; социальная организация, основанная на родстве по материнской линии. Эта классификация поведенческих особенностей создает подходящую основу для формирования гипотез. Она позволяет качественно оценить вероятность того, что различные поведенческие особенности сохранились у современного Homo sapiens. Конечно, сохраняется возможность того, что некоторые лабильные черты гомологичны у человека и, скажем, шимпанзе. И наоборот, некоторые черты, консервативные у всей остальной части приматов, могли претерпеть изменения по мере происхождения человека. Кроме того, такая оценка не подразумевает, что консервативные черты в большей мере определяются генетически, т.е. имеют более высокую наследуемость, чем лабильные. Лабильность может быть основана полностью на генетических различиях между видами или видовыми популяциями. Возвращаясь, наконец, к вопросу о культурной эволюции, мы можем эвристически догадываться, что отличия между человеческими сообществами по характеристикам, аргументировано отнесенным к лабильным, наиболее вероятно определяются генетическими различиями. Свидетельства, рассмотренные в Таблице 26-1, не противоречат этой основной концепции. Наконец, и это важно заметить особо, что сравнительно-этологический подход не всегда способен прогнозировать уникальные особенности человека. Общее правило эволюционного исследования в том, что направление значительных “скачков” не так легко читается с помощью филогенетической экстраполяции.
Таблица 26-1. Основные социальные характеристики человека, расположенные в соответствии с тем, являются ли они уникальными, лабильными (изменчивыми на видовом/родовом уровне у остальных приматов) или консервативными (однотипными у всех приматов).
| Эволюционно лабильные признаки приматов |
Эволюционно консервативные признаки приматов |
Человеческие признаки |
| ОБЩИЕ С НЕКОТОРЫМИ ДРУГИМИ ПРИМАТАМИ |
||
| Размер группы |
Высоко изменчив |
|
| Сплоченность группы |
Высоко изменчива |
|
| Открытость группы для других |
Высоко изменчива |
|
| Участие самца в заботе о потомстве |
Существенное |
|
| Структура внимания |
Центростремительная, направлена на лидирующих самцов |
|
| Интенсивность и форма защиты территории |
Высоко изменчивы, но территориальность общая |
|
| ОБЩИЕ СО ВСЕМИ ИЛИ ПОЧТИ СО ВСЕМИ ДРУГИМИ ПРИМАТАМИ |
||
| Системы агрессивного доминирования, где самцы доминируют над самками |
Соответствуют таковым у других приматов, хотя и изменчивы |
|
| Шкалирование реакций, особенно при агрессивных взаимодействиях |
Соответствуют таковому у других приматов |
|
| Длительная материнская забота, выраженная социализация молодых |
Соответствуют таковым у других приматов |
|
| Организация, основанная на родстве по материнской линии |
В основном соответствует таковой у других приматов |
|
| УНИКАЛЬНЫЕ |
||
| Речь, сложная культура |
||
| Половая активность непрерывна в течение менструального цикла |
||
| Формальные инцест-табу, правила брачных обменов с распознаванием отношений родства |
||
| Кооперативное разделение труда между взрослыми мужчинами и женщинами |
||
Товарообмен и реципрокный альтруизм
Обмены среди приматов редки, существуя в зачаточной форме только у шимпанзе и, возможно, у некоторых других обезьян Старого Света. Однако у человека это одна из самых сильных социальных черт, достигающая уровня, сопоставимого с обменом пищей у термитов и муравьев. В результате, только человек создал экономику. Его высокий интеллект и способность к символизации делают возможным истинный бартер. Кроме того, интеллект позволил, чтобы обмены были отсроченными, превратив их в акты реципрокного альтруизма (Trivers, 1971). Традиции этого способа поведения выражены в знакомых высказываниях из каждодневной жизни:
“Дай мне что-нибудь сейчас, а я расплачусь потом”.
“Помоги мне сегодня, и ты сможешь рассчитывать на мою дружбу, когда я тебе понадоблюсь”.
“Я действительно не думал о спасении как о героизме, я делал только то, чего жду от других в подобной ситуации для себя и своей семьи”.
Деньги, как любил подчеркивать Тэлкотт Парсонс, не имеют никакого значения сами по себе. Это только кусочки металла и клочки бумаги, которыми люди довольствуются, отказываясь в пользу другого от прав собственности на различное количество имущества или услуг по требованию. Другими словами, это количественное выражение реципрокного альтруизма.
Возможно, самой первой формой бартера в ранних человеческих обществах был обмен мясом, добытым мужчинами, на растительную пищу, собранную женщинами. Если существующие общества охотников и собирателей отражают примитивное состояние, этот обмен образовывал важный элемент в особом виде связи между полами.
Фокс (Fox, 1972), вослед Леви-Страуссу (Lévi-Strauss, 1949), исходя из этнографических данных, привел доводы, что ранним и ключевым шагом человеческой социальной эволюции было использование женщин в бартере. Так как мужчины приобретали статус благодаря контролю над женщинами, они использовали их как объект обмена для скрепления альянсов и поддержания родственных связей. Дописьменные общества характеризуются сложными брачными правилами, которые можно интерпретировать буквально как гонорар за могущество. В частности, это имеет место, когда элементарные запретительные правила брака, отвергающие некоторые типы союзов, сочетаются с разрешительными правилами, устанавливающими, какие именно обмены должны быть сделаны. В австралийских аборигенных сообществах племена разделены на две половины, между которыми разрешены браки. Мужчины каждой такой половины торгуют племянницами, точнее, дочерьми своих сестер. Могущество крепнет с возрастом, поскольку мужчина может контролировать и потомков племянниц, вплоть до дочерей дочери его сестры. Вкупе с многоженством, эта система обеспечивает старикам племени как политическое, так и генетическое преимущество.
При всей ее запутанности, формализация брачных обменов между племенами имеет приблизительно тот же генетический эффект, как и случайное блуждание самцов обезьян от одной группы к другой или обмен молодыми зрелыми самками между популяциями шимпанзе. Приблизительно 7,5% браков, заключенных между аборигенами Австралии до европейского влияния, были межплеменными. Подобный же уровень был зафиксирован у бразильских индейцев и других дописьменных обществ (Morton, 1969). Элементарная теория популяционной генетики говорит, что поток генов порядка 10% на поколение более чем достаточен для противодействия довольно интенсивному давлению естественного отбора, стремящемуся дифференцировать популяции. Таким образом, межплеменные брачные обмены являются ведущим фактором поддержания наблюдаемой высокой степени межпопуляционного генетического подобия. Решающее же адаптивное значение экзогамии не в потоке генов как таковом, а, скорее, в предотвращении инбридинга. И вновь, 10% потока генов адекватны этой цели.
Микроструктура социальной организации людей основана на сложных взаимных оценках, предшествующих заключению контрактов. Как правильно уловил Эрвин Гофман, чужака быстро, но деликатно исследуют, чтобы определить его социально-экономический статус, интеллект и образование, самооценку, социальные отношения, компетентность, кредитоспособность и эмоциональную стабильность. Эта информация, во многом предоставляемая и воспринимаемая подсознательно, имеет чрезвычайно важное практическое значение. Зондирование должно быть глубоким, ибо индивидуум пытается создать впечатление, которое обеспечит ему максимальные преимущества. По крайней мере, он маневрирует, чтобы не открыть информацию, которая подвергнет опасности его статус. Можно заранее ожидать, что представление самого себя будет содержать элементы, вводящие в заблуждение.
Много важных обстоятельств остаются вне времени и места взаимодействия или скрываются. Например, достоверные или “истинные” отношения, мнения и эмоции индивидуума можно определить только косвенно, через его нечаянные признания или через то, что выглядит как невольно проявляющееся экспрессивное поведение. Если кто-то предлагает другим продукт или услугу, те часто оказываются перед фактом, что в ходе общения у них не будет возможности здесь и сейчас отведать сего пудинга, чтобы получить подтверждение его качества. Они будут вынуждены воспринять некоторые вещи как обычные или естественные признаки чего-то, непосредственно недоступного чувствам (Goffman, 1959).
Обман и лицемерие не являются ни абсолютными грехами, которые добродетельные люди сводят к минимуму, ни остаточными животными чертами, которым предстоит быть стертыми дальнейшей социальной эволюцией. Это специфические приспособления человека для устройства сложных ежедневных дел социальной жизни. В каждом конкретном сообществе их уровень представляет компромисс, отражающий размер и сложность данного сообщества. Если уровень слишком низок, другие завладеют преимуществом и победят. Если слишком высок, – грозит изгнание из общества. Полная честность, как ни посмотри, – не выход. Прежнее “простодушие” приматов уничтожило бы тонкую ткань социальной жизни, возросшей в человеческих популяциях за пределами кланов. Как справедливо подметил Луис Дж. Халл, хорошие манеры заменили нам любовь.
Сексуальные и другие связи
Кирпичиком почти всех человеческих обществ является семейная ячейка (Reynolds, 1968; Leibowitz, 1968). Население индустриального американского города в той же мере, как и группа охотников и собирателей в австралийской пустыне, организовано вокруг этой единицы. В обоих случаях семья перемещается между региональными общинами, поддерживая сложные связи с непосредственными родственниками с помощью визитов (или телефонных звонков и писем) и обмена подарками. В течение дня женщины и дети остаются в местах постоянного проживания, в то время как мужчины добывают дичь или ее символический эквивалент в форме предметов для обмена и денег. Мужчины объединяются в группы для охоты или взаимодействия с соседствующими группами. И даже если группу не объединяет кровное родство, она имеют склонность действовать, по меньшей мере, как “банда братков”. Половые связи обусловлены тщательным соблюдением обычаев племени и предполагают долговременность. Полигамию, как тайную, так и явную – одобренную обычаями, практикуют преимущественно мужчины. Половое поведение возможно в течение всего женского менструального цикла и отличается длительной предшествующей игрой. Моррис (1967), используя данные Мастерса и Джонсон (Masters and Johnson, 1966) и других, перечисляет уникальные признаки человеческой сексуальности, которые, по его мнению, связаны с потерей волос на теле: округлые и выдающиеся вперед груди молодых женщин; покраснение участков кожи во время совокупления; расширение сосудов и повышенная эрогенная чувствительность губ, мягких частей носа, ушей, сосков, околососковых кружков и гениталий; большой размер пениса у мужчин, особенно при эрекции. Как заметил сам Дарвин в 1871 году, даже обнаженная кожа женщины используется как сексуальный релизер. Все эти изменения служат “цементированию” постоянных связей – вне зависимости от времени наступления овуляции. Проявление эструса уменьшилось до рудиментарного – к разочарованию тех, кто пытается практиковать в контрацепции метод “естественного цикла”. Половое поведение стало мало связанным собственно с оплодотворением. По иронии судьбы, святоши, порицая занятие сексом с иной целью, чем произведение потомства, ссылаются на “законы природы”. Однако это – следствие ошибочного вывода сравнительной этологии, основанного на неправильном предположении о том, что в размножении человек подобен, по существу, другим животным.
Расширение и формализация родственных связей, господствующие почти во всех человеческих обществах, – тоже уникальная особенность биологии нашего вида. Системы родства обеспечивают, по меньшей мере, три явных преимущества. Во-первых, они способствуют альянсам между племенами и внутриплеменными единицами, что обеспечивает канал для бесконфликтной эмиграции молодых членов. Во-вторых, они являются важной частью систем меновой торговли, с помощью которой определенные мужчины достигают лидирующего положения и доминирования. Наконец, они служат гомеостатическим устройством, позволяющим группе переживать трудные времена. Когда пищи становится недостаточно, племенные единицы могут обратиться к союзникам за альтруистической помощью, что совершенно неизвестно у других общественных приматов. Индейцы атапаски догриб – племя охотников и собирателей, проживающих в Арктике на северо-западе Канады, дают нам первый пример. Атапаски организованы не жестко, по принципу двусторонних элементарных связей (June Helm, 1968). Локальные группы перемещаются по общей территории, периодически контактируя и обмениваясь членами для родственных браков. Когда случается голод, находящиеся под угрозой вымирания группы могут присоединяться к тем, которые в данный момент благополучнее их. Второй пример – Яаномамо в Южной Америке, полагающиеся на родню, когда посевы истреблены врагами (Chagnon, 1968).
Поскольку общества прошли путь развития от первобытного человеческого стада через племена к тираниям и государствам, некоторые типы связей были расширены за пределы родственных сетей и включили в свой состав другие типы альянсов и экономических соглашений. Поскольку эти сети стали больше, линии сообщения длиннее, взаимодействия разнообразнее, система в целом стала в высшей степени сложной. Но нравственные законы, лежащие в основе общественных структур, по всей видимости, не слишком менялись. Средний гражданин по-прежнему действует в рамках формализованного кодекса, не более совершенного, чем тот, который управляет членами обществ охотников и собирателей.
Исполнение ролей и разделение труда
Супермен, как и супер-муравей или супер-волк, не могут в принципе существовать в одиночку – только в группе, разнохарактерные члены которой совместно составляют общество, возможности которого превосходят таковые любого мыслимого отдельного организма. Человеческие общества вознеслись до уровня чрезвычайной сложности благодаря интеллекту и гибкости своих членов, которые могут играть поистине любую роль, необходимую в данных обстоятельствах, а если нужно, то и переключаться между ними. Современный человек – актёр многих амплуа, который хорошо умеет выходить за пределы своих возможностей в соответствии с постоянно меняющимися требованиями внешней среды. Как отметил Гоффман (Goffman, 1961), “возможно, временами человек марширует взад-вперед подобно оловянному солдатику, закованный в мундир исполняемой роли. Действительно, иногда мы можем уловить тот момент, когда он полностью “оседлывает” единственную роль (голова поднята, отрешенный взгляд), но уже в следующий момент эта картина рассыпается и личность разбивается на различные персонажи, удерживающие звенья различных сфер жизни с помощью своих рук, своих зубов, своей мимики. При ближайшем рассмотрении индивидуум, всеми способами удерживающий имеющиеся в его в жизни взаимоотношения, просто теряет очертания”. Ничего удивительного в том, что наиболее острой внутренней проблемой современного человека является проблема идентичности.
Роли в человеческих обществах принципиально отличаются от каст социальных насекомых. Члены человеческих обществ иногда тесно сотрудничают, подобно социальным насекомым, но чаще они конкурируют за обладание ограниченными ресурсами, локализованными в их ролевой сфере. Наилучшие и наиболее предприимчивые из исполняющих роли актеров обычно получают непропорциональную долю вознаграждения, при этом наименее успешные отодвинуты на другие, менее приятные, позиции. Кроме того, индивидуумы пытаются повысить свое социально-экономическое положение, меняя роли. Происходит также и соревнование между классами, и в переломные моменты истории это предопределяет общественные перемены.
Ключевой вопрос биологии человека: предопределены ли генетически принадлежность к определенному классу и амплуа жизненной роли? Можно легко себе представить обстоятельства, при которых могла бы происходить такая генетическая дифференциация. Передачи по наследству по крайней мере некоторых параметров интеллекта и эмоциональных особенностей достаточно для соответствия умеренному дизруптивному отбору. Дальберг (Dahlberg, 1947) показал, что если возникает единственный ген, ответственный за успех и возвышение в обществе, он может быть быстро сконцентрирован в высших социально-экономических классах. Предположим, например, что имеются два класса, начавших каждый только с 1% частоты гомозигот по стремящемуся наверх гену. Предположим далее, что в каждом поколении 50% гомозигот из низшего класса перемещаются в высший. Тогда уже в десятом поколении, в зависимости от относительных размеров групп, высший класс будет обладать 20% гомозигот или даже больше, а низший – только 0.5% или меньше. Используя сходный аргумент, Геррнштейн (Herrnstein, 1971b) предположил, что поскольку возможности, определяемые средой, стали почти равным в пределах обществ, разделение на социально-экономические группы будет все более и более определяться генетическими различиями в интеллекте.
Сильный начальный сдвиг в сторону такого расслоения возникает, когда одно человеческая популяция побеждает и порабощает другую – достаточно обычный случай в человеческой истории. Генетические различия в умственных способностях, как бы они ни были малы, стремятся быть сохраненными посредством возведения классовых барьеров, расовой и культурной дискриминации, и физически – с помощью гетто. Генетик Дарлингтон (К.D. Darlington, 1969), среди прочих, предположил, что этот процесс может быть главным источником генетического разнообразия в человеческих обществах.
Все же, несмотря на правдоподобие общих рассуждений, имеется мало свидетельств в пользу какой-либо передачи по наследству устоявшегося статуса. Касты Индии существуют уже в течение 2000 лет, это более чем достаточное время для эволюционной дивергенции, но касты лишь слегка отличаются по типам крови и другим измеримым анатомическим и физиологическим характеристикам. Можно указать на мощные силы, работающие против генетической фиксации кастовых различий. Во-первых, культурная эволюция слишком нестабильна. За десятки лет, максимум – сотни, гетто вытеснены, расы и народы освобождены от плена, завоеватели усмирены. Даже в относительно устойчивых обществах есть многочисленные тропинки наверх. Дщери низших классов стремятся выходить замуж “с повышением”, за мужчин более высокого класса. Успех в коммерческой или политической жизни может перебросить семейство фактически из любой социально-экономической группы в правящий класс за одно-единственное поколение. Более того, имеется много “генов Дальберга”, а не тот один, который мы рассматривали в простейшей модели. Наследственные факторы человеческого успеха чрезвычайно полигенны и образуют длинный список, но лишь малая часть из них изучена. КИ (“коэффициент интеллектуальности”, или “коэффициент умственного развития” – прим. перев. М.П.) – только одно подмножество из многих компонентов интеллекта. Менее доступными измерению, но не менее важными качествами являются творческий потенциал, предприимчивость, настойчивость в достижении цели и умственная работоспособность. Допустим, что поддерживающие эти качества гены рассеяны по многим хромосомам. Предположим также, что некоторые из этих особенностей не коррелируют или даже отрицательно коррелируют друг с другом. При этих обстоятельствах лишь наиболее сильные формы дисруптивного отбора могли привести к формированию устойчивых совокупностей генов. Более правдоподобной выглядит вот какая, по всей видимости, преобладающая ситуация: поддерживается широкое генетическое разнообразие в пределах обществ при слабой связи некоторых генетически обусловленных характеристик с собственно успехом. Этот сложный процесс ускоряется постоянными изменениями в судьбах отдельных семейств от одного поколения к следующему.
Даже в этом случае нельзя игнорировать влияние генетических факторов на усвоение конкретных и отчетливых ролей. Рассмотрим мужской гомосексуализм. Исследования Кинси (Kinsey) и его коллег показали, что в 1940-х гг. приблизительно 10% от достигших сексуальной зрелости мужчин в Соединенных Штатах были главным образом или исключительно гомосексуалистами, по крайней мере, в течение трех лет до опроса (более поздние исследования эту цифру не подтвердили. В 1990-х гг. гомосексуалистов в США насчитали не более 3%. Или времена изменились, или один из опросов был неточен – прим. перев. А.П.). Гомосексуализм также в сравнительно высоких долях присутствует среди мужского населения во многих, если не в большинстве, остальных культурах. Проведенное на близнецах исследование Каллманна (Kallmann) свидетельствует о вероятном существовании генетического предрасположения к этому состоянию. Соответственно, Хатчинсон (Hutchinson, 1959) высказал предположение, что гены гомосексуальности в гетерозиготном состоянии могут придавать их носителю ряд очень полезных качеств. Эта аргументация нашла своих последователей, теперь это – общее место в популяционной генетике. Следствием гомосексуального статуса как такового является низкая генетическая продуктивность подобной особи, ведь люди нетрадиционной ориентации женятся гораздо реже и имеют гораздо меньше детей, чем их однозначно гетеросексуальные коллеги. Простейшим способом эволюционного сохранения генов, вызывающих такое состояние, может быть их преимущество в гетерозиготном положении: гетерозиготы лучше доживают до зрелости и/или производят больше потомков. Интересная альтернативная гипотеза была предложена Германом Т. Спитом (Herman T. Spieth, личное сообщение). Гомосексуальные члены примитивных обществ могли выступать помощниками в компании с другими мужчинами на охоте, либо участвовать в домашних делах в жилищах. Будучи свободны от выполнения родительских обязанностей, они могли быть особенно полезны в помощи близким родственникам. Гены предрасположенности к гомосексуализму могли, таким образом, равновесно поддерживаться на достаточно высоком уровне за счёт одного лишь родственного отбора. Остается сказать, что если такие гены действительно существуют, они почти наверняка обладают неполной пенетрантностью (проявляются не у всех их носителей – прим. перев. М.П.) и изменчивы по экспрессивности (проявляются в разной степени – прим. перев. М.П.), что означает, что проявят ли и в какой степени проявят носители генов соответствующие поведенческие особенности зависит от присутствия или отсутствия генов-модификаторов и воздействия окружающей среды. Недавний анализ этнографических данных (Weinrich, 1976) показал, что гомосексуалисты в современных сообществах охотников и собирателей действительно благотворно влияют на родственников, зачастую выступая в роли шаманов и бердачей (“мужеженщин” – людей с недифференцированным или противоположным биологическому гендерным статусом – прим. перев. А.П.).
Возможно, существуют и другие основные типы, и весьма вероятно, что ключ к разгадке будет найден только при полном изучении вопроса. В своем исследовании, проведенном в детских учреждениях Британии, Блартон Джонс (Blurton Jones, 1969) выделил у детей две основные модели поведения, в соответствии с которыми их можно разделить на “тех, кто говорит” и на “тех, кто делает” (или “вербалистов” и “мастеров” в соответствии с психологической терминологией – прим. перев. С.Р.). Первые, составлявшие меньшинство, часто пребывали в одиночестве, редко передвигались с места на место, и почти никогда не участвовали в шумных забавах. Они были словоохотливыми рассказчиками и проводили много времени за чтением книг. Вторые объединялись в группы, много двигались и предпочитали больше рисовать и мастерить, чем тратить время на разговоры. Блартон Джонс предположил, что эта дихотомия является следствием ранней дивергенции в развитии поведения, сохраняющейся потом у человека и в зрелые годы. В общем случае это может основополагающим образом влиять на многообразие внутри культур. Пока неизвестно, предопределено ли это расхождение путей развития только генетически, или оно полностью обусловлено эмпирическим опытом раннего детства.
Общение
Все уникальное социальное поведение человека основывается на его способности пользоваться речью, которая, в свою очередь, тоже уникальна. В любом языке слова, которым присвоены произвольные определения в рамках культур, выстроены в соответствии с грамматикой, которая придает им новые оттенки значений за пределами основного смысла. Полная символичность слов и изощренность грамматик позволяет создавать, в принципе, неограниченное множество сообщений. Можно даже общаться на тему самой системы общения! В этом – сущность человеческого языка. Базовые атрибуты общения могут быть разрушены, и другие особенности передачи информации могут быть добавлены, а в итоге формируются 16 основополагающих признаков (C. F. Hockett, см. в обзоре: Thorpe, 1972a). Большинство этих признаков найдено, как минимум, в рудиментарной форме и у других видов животных. Но продуктивности и богатства человеческого языка не может достичь никто, даже шимпанзе, обученные использованию знаков в простых предложениях. Развитие человеческой речи представляет собой эволюционный “квантовый скачок”, сравнимый с объединением эукариотических клеток в многоклеточные организмы.
Человек способен к богатейшему общению даже без слов. Изучение невербальной коммуникации ныне – процветающая ветвь социальных наук. Её кодификация затруднена тем, что очень многие сигналы играют вспомогательную роль и в вербальном общении. Категории этих сигналов зачастую определены противоречиво, и классификации редко согласуются (см., напр., Rensky, 1966; Crystal, 1969; Lyons, 1972). В таблице 26-2 представлена суммарная схема, свободная, как мне кажется, от внутренних противоречий и согласующаяся с текущим использованием. Число неголосовых сигналов, включая выражения лица, положения тела, его движения и прикосновения, вероятно, несколько превышает цифру 100. Брэнниган и Хамфрис (Brannigan and Humphries, 1972) составили список из 136 таких сигналов, который, как они считают, вполне исчерпывающ. Это число согласуется с полностью независимой оценкой Бердвистла (Birdwhistle, 1970), который полагает, что хотя лицо человека способно отобразить 250 тысяч выражений, однако число уникальных осмысленных значений из них не более 100. Вокальный параязык, насколько его можно отделить от ритмико-интонационных модификаций настоящей речи, не был каталогизирован так же досконально. Грант (Grant, 1969) распознал 6 уникальных звуков, но нередко это число подвергается сомнению зоологами, привыкшими препарировать этограммы других видов приматов. Короче говоря, всех паралингвистических сигналов, вместе взятых, почти наверняка больше 150, и, может быть, около 200. Этот запас больше, чем у основной массы других млекопитающих и птиц раза в три или больше, и несколько превосходит репертуар макак-резусов и шимпанзе.
Таблица 26-2. Способы общения людей.
|
I. Вербальная коммуникация (язык): произнесение слов и предложений II. Невербальная коммуникация А. Интонации: тон, темп, ритм, громкость, интервал и другие характеристики голоса, модифицирующие значение произнесённых слов Б. Пара-язык: сигналы, отличные от слов, используемые для дополнения или модификации речи 1. Вокальный параязык: мычание, хихиканье, смех, крики и другие невербальные звуки 2. Невокальный параязык: положение тела, движение и прикосновения (кинестическая коммуникация); возможно также химическая коммуникация |
Другое важное отличие в анализе человеческого параязыка может быть отмечено между прелингвистическими сигналами, определенными как эволюционные предтечи настоящего языка, и теми, что являются постлингвистическими. Постлингвистические сигналы, вероятнее всего, произошли как чисто вспомогательные для речи. Ван Хуфф (Van Hooff, 1972), к примеру, установил гомологичность улыбки и смеха в выражениях лица низших и высших обезьян Старого Света, таким образом, классифицировав эти поведенческие акты как наиболее древние и универсальные сигналы.
Как доказывал Марлер (Marler, 1965), человеческая речь, возможно, основывается на богатстве и разнообразии сигналов, которые вряд ли отличны от тех, что использовались макаками-резусами или шимпанзе, в противоположность более разрозненным сигналам, характеризующим репертуар низших приматов. Человеческие младенцы могут издавать широкий спектр вокализаций, похожий на звуки, издаваемые макаками, павианами, и шимпанзе. Но очень рано в ходе их развития такие звуки преобразуются в специфические звуки человеческой речи. Многочисленные взрывные, фрикативные, носовые, гласные и другие звуки комбинируются, чтобы породить 40 или около того, основных фонем. Рот человека и верхние дыхательные пути серьезно модифицированы, чтобы поддержать эти вокальные навыки (см. рис. 26-2). Решающие изменения ассоциированы с прямохождением человека, которое могло быть обеспечено начальным, но все еще не закончившимся импульсом в направлении нынешней модификации. На лице, обращенном полностью вперед, рот образует с верхней гортанью угол 90 градусов. Такая конфигурация помогает поднимать заднюю часть языка, пока не получится единый тракт с частью передней стенки верхней гортани. Одновременно область гортани и надгортанник намного удлиняются.
|
|
|
Рисунок 26-2. Голосовой аппарат человека модифицировался в ходе развития так, чтобы значительно расширить множество издаваемых звуков. Подвижность была существенным аккомпанементом в развитии речи человека. Верхняя диаграмма показывает отличия вокального аппарата человека от шимпанзе и других приматов. Угол между ртом и верхним дыхательным трактом увеличен, область гортани удлинена, а задняя половина языка приобрела форму передней стенки длинного тракта над голосовыми связками. Нижняя диаграмма иллюстрирует, как перемещения языка изменяет резонирующие объемы и порождает различные звуки (приводится с изменениями по: Howells, 1973; Denes and Pinson, 1973). |
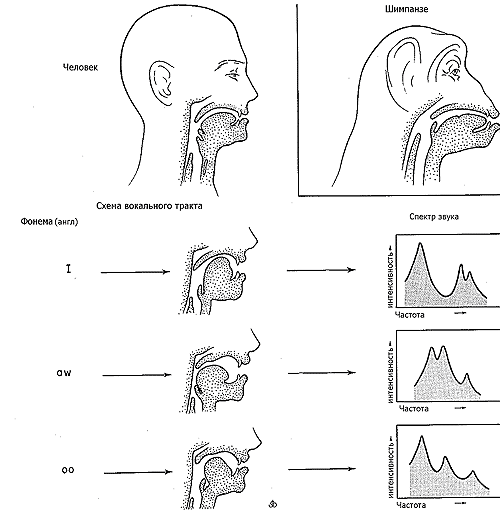
Два принципиальных измерения – переменчивость положения языка, и удлинение гортани ответственны за многообразие издаваемых звуков. Когда воздух с силой проходит вверх сквозь голосовые связки, он производит жужжащий шум, который может быть сильнее или продолжительнее, но не может претерпеть все важные модификации тона, чтобы произвести дифференциацию фонем. Последние эффекты достигаются при прохождении воздуха через гортанные и ротовые полости, и через ротовую щель. Эти структуры совместно формируют духовую трубу, которая, как и любой цилиндр, служит резонатором. Когда его расположение или очертания меняются, труба акцентирует различные комбинации частот, производимых голосовыми связками (голосовые связки, будучи колебательной системой с низкой добротностью, генерируют звук с широким спектром гармоник, т.е. частот, сопутствующих основному тону; именно эти гармоники и выделяют описанные резонаторы. Если бы голосовые связки порождали чистый тон, резонаторы любого типа влияли бы лишь на громкость – прим. перев. А.П.). В итоге, как показано на рис 26-2, мы различаем звуки как фонемы (см. также: Lenneberg, 1967; Denes and Pinson, 1973).
Однако наиболее ценное свойство дара речи – это не столько способность производить много звуков. В конце концов, теоретически возможно, что очень умное существо, произносящее единственное слово, будет способно к интенсивному общению. Необходимо лишь быть запрограммированным, как цифровой компьютер. Оно могло бы быть дополнено вариациями громкости, длительности, и периода следования, и тем еще более увеличить информативность коммуникации. В самом деле, единственное химическое вещество, в случае модулирования в идеальных условиях может производить до 10 000 бит в секунду, что далеко превосходит информативность человеческой речи. Зато человеческая речь сильна своим синтаксисом, зависимостью значений от порядка следования слов. Каждый язык обладает грамматикой, набором правил, управляющих синтаксисом. Чтобы верно понять природу и происхождение грамматики, нужно узнать почти все о конструкции человеческого рассудка. Можно отметить три конкурирующие модели, пытающиеся описать известные правила:
Первая гипотеза: вероятностная модель “слева–направо”. Излюбленное объяснение крайних психологов-бихевористов в том, что встречаемость слова подчиняется марковскому распределению, т.е. его вероятность зависит от непосредственно предшествующих слов или групп слов. Развивающийся ребенок усваивает эти слова во взаимосвязи с каждой подобающей обстановкой.
Вторая гипотеза: модель усвоенной глубокой структуры. Имеется ограниченный набор формальных принципов, в соответствии с которым слова комбинируются и составляются во фразы, порождая различные значения. Ребёнок в той или иной степени неосознанно изучает глубокую структуру его собственной культуры. Хотя число этих принципов ограничено, количество предложений, которые могут порождаться в соответствии с ними, неограниченно. Животные не могут говорить просто потому, что у них нет необходимого уровня познавательных или мыслительных способностей, но не потому, что у них нет какой-то специфической “способности к речи”.
Третья гипотеза: модель врожденной глубокой структуры. Некие формальные принципы существуют, как и во второй гипотезе, но они частично или полностью наследственные. Другими словами, по меньшей мере, некоторые из принципов инвариантно появляются в ходе созревания. Естественное следствие из этого предположения в том, что большая часть глубокой структуры грамматики общераспространенна, если не универсальна, во всём человечестве, хотя существуют глубокие различия во внешней структуре и значении слов между языками. Второй естественный вывод в том, что животные не могут говорить потому, что у них нет этой врождённой способности к речи, которая является качественно уникальной особенностью человека, но не является результатом количественного превосходства в интеллекте. Гипотеза врожденной глубокой структуры ярко ассоциируется с именем Ноэма Хомски (Noam Chomsky), и кажется сейчас в фаворе у большинства психолингвистов.
Вероятностная модель “слева–направо” сейчас отвергнута, по крайней мере – в её экстремальной версии. Количество переходных вероятностей, которые ребёнок должен изучить, чтобы вычислить такой язык как английский – практически беспредельно, и у него просто не хватит времени, чтобы в течение детства освоить все их (Miller, Galanter, and Pribram, 1960). На практике грамматические правила осваиваются очень быстро и в предикативных предложениях, ребенок проходит конструкции языка, предвосхищающие взрослые формы, одновременно различая важность их (Brown, 1973). Такой ход онтогенеза типичен для созревания врожденных компонент поведения животных. Тем не менее, это сходство не может приниматься как решающее свидетельство в пользу генетической программы, общей для человечества.
Окончательного решения этой проблемы, как настаивает Роджер Браун (Roger Brown) и другие представители психолингвистики развития, невозможно достичь, пока глубокая грамматика не будет надежно охарактеризована. Это сравнительно новая область исследования, начавшаяся с книги Хомски “Синтаксические структуры” (Syntactic Structures, Chomsky, 1957). С самого начала она отмечена высокосложной и быстроменяющейся аргументацией. Основные идеи были представлены в обзорах (Slobin, 1971; Chomsky, 1972). Здесь достаточно будет охарактеризовать главные процессы, распознанные в новом лингвистическом анализе. Грамматика структуры фраз, которая послужила примером в рис 26-3, содержит иерархию правил, по которым строятся предложения. “Фразы” (“выражения”), могут рассматриваться как модули, которые заменяют другие, эквивалентные, модули или добавляются de novo в предложение, чтобы изменить значения (обращаю внимание, что здесь и чуть далее “фраза” понимается не как синоним грамматического предложения, а более узко и строго – как некая единица речи – прим. перев. А.П.). Эти элементы нельзя разорвать, и части нельзя, без серьезных затруднений, менять местами. В примере “Мальчик ударил по мячу” “по мячу” интуитивно понимается как некая единица, которая легко может быть удалена или заменена другим выражением, к примеру, “по волану”, или просто “по нему”. Комбинация “ударил по” – уже не такая единица. Несмотря на тот факт, что эти два слова сопоставлены, их нельзя легко заменить без затруднений в конструировании остальной части предложения. Соблюдая правила, которые мы все подсознательно знаем, мы можем расширить предложение вставкой соответственно выбранных фраз: “Заняв свою позицию, маленький мальчик дважды замахнулся и, наконец, ударил по мячу и побежал к первой базе” (похоже, речь о бейсболе – прим. перев. М.П.).
|
|
|
Рисунок 26-3. Пример структуры правил грамматики в английском языке. Простое предложение “Мальчик ударил по мячу” (“The boy hit the ball”) показано как содержащее иерархию фраз. Каждый уровень одной фразы можно заменить другой эквивалентной композицией, но фразы нельзя расщеплять и менять местами их элементы (на основе: Slobin, 1971). |
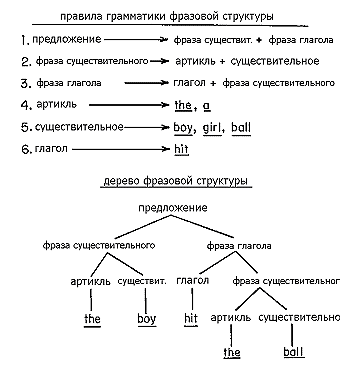
Короче говоря, грамматика фразовой структуры предписывает способы формирования фраз. Они производят то, что называется глубокой структурой групп слов, в противоположность наружной структуре, или не более чем порядок, в котором появляются отдельные слова. Но направление последовательности, в которой фразы и терминальные слова появляются имеет решающее значение для смысла предложения. Предложение “Мальчик ударил по мячу” очень отличается от “По чему ударил мальчик?”, несмотря даже на то, что глубокая (фразовая) структура этих предложений подобна. Правила, в соответствии с которыми глубокая структура преобразуется в наружную структуру посредством сборки фраз, называется “преобразовательной грамматикой”. Преобразование – это операция конвертирования одной фразовой структуры в другую. Среди основных операций – замены (“по чему” заменяется на “по мячу”), перестановки (“по чему” ставится перед глаголом), и перемещения (изменение расположения связанных слов).
Психолингвисты описали как фразовую структуру, так и преобразовательную грамматику английского языка. Однако их аргументация не выглядят достаточной, чтобы выбрать между гипотезами номер два и три; другими словами – чтобы решить, являются ли грамматики унаследованными программами, или им обучаются. Базовые операции преобразования существуют во всех известных человеческих языках. Однако это наблюдение само по себе не позволяет утверждать, что конкретные правила преобразований одинаковы.
Существует ли универсальная грамматика? На этот вопрос ответить трудно в силу того, что большинство попыток обобщить правила глубокой грамматики базировались на семантическом содержании некоего отдельно взятого языка. Изучающие этот предмет редко стоят перед проблемой как истинно научной, хотя лишь этот путь мог бы быть привести к успеху. В самом деле, ученых-естественников разочаровывают расплывчатость и косвенность, которыми грешит большая часть психолингвистической литературы, которая выглядит зачастую легкомысленной и не заботящейся о классических канонах подведения доказательной базы под свои утверждения. Причина в том, что многие авторы, включая Хомски, – структуралисты в духе Леви-Страусса и Пиаже. Они подходят к проблеме с четким воззрением на человеческий разум (практически – мировоззрением), что он (разум) тоже структурирован, и тоже дискретен, неисчислим, эволюционно уникален, и, следовательно, нет большой нужды описывать его в формулировках других научных дисциплин. Анализ этот не теоретический, в смысле, что его нельзя вывести из проверяемых опытом постулатов, и расширить эмпирически. Некоторые психологи, включая Роджера Брауна с коллегами, а также Фодора и Гарретта (Fodor and Garrett, 1966), выдвигали и исследовали с переменным успехом поддающиеся проверке предположения, но следить за спекуляциями о глубокой грамматике было нелегко даже для этих искушенных экспериментаторов.
Подобно поэтам-натуралистам, структуралисты воспевают идиосинкразическое уникальное персональное видение. Их аргументация исходит из скрытых допущений, они весьма полагаются на метафоры и поясняющие примеры, уделяя мало внимания методу множественных конкурирующих гипотез. Несомненно, что эта дисциплина, одна из наиболее важных среди всех наук, созрела для применения строгих теорий и должной связи с экспериментальными исследованиями.
Фокус в том, что новая лингвистика может никогда не ответить на вопрос о том, когда у человека появилась речь. Появилась ли она с первым использованным каменным рубилом и постройкой жилищ австралопитеками более 2 миллионов лет назад? Или она ждала появления полностью современного Homo sapiens, или, может, даже развития религиозных обрядов в последние 100 тысяч лет? Либерман (Lieberman, 1968) полагает, что этот момент был относительно недавно. Он изучил реконструкции Макапанских австралопитеков, сделанные Дартом, и нашел, что форма их нёба и гортани очень близка к шимпанзе. Если он прав, то это значит, что ранние гоминиды не были способны артикулировать звуки человеческой речи. Такие же выводы можно сделать из изучения анатомии и вокальных способностей неандертальского человека (Lieberman et al., 1972), и если это верно, то точка возникновения человеческой речи находится на позднейших этапах видообразования в роде Homo. Другой теоретический аспект эволюционного происхождения человеческой речи был обсужден Джейн Хилл (Jane Hill, 1972) и Мэттингли (I. G. Mattingly, 1972). Леннеберг (Lenneberg, 1971) выдвинул гипотезу, что способность к математическим рассуждениям возникла как незначительная модификация лингвистических способностей.
Культура, ритуалы и религия
Зачатки культуры имеются, кроме человека, и у других высших приматов, в том числе, у японской макаки и у шимпанзе; но только у человека культура получила настолько полное развитие, что ею пронизаны буквально все аспекты его жизнедеятельности. Этнографические характеристики генетически прописаны нечетко, результатом чего является огромное разнообразие описываемых сообществ. Недостаточная определенность не означает, что культура совершенно не зависит от генетических характеристик и свойств. Развивалась собственно способность к культуре; в действительности можно говорить о присущей обществу упорной тенденции к формированию культуры в той или иной возможной форме. Робин Фокс (Robin Fox, 1971) излагает этот аргумент в следующем виде. Если бы эксперименты фараона Псамметиха и короля Шотландии Джеймса IV увенчались успехом и воспитываемые в изоляции дети каким-то образом выросли здоровыми,
я не сомневаюсь в том, что они могли бы говорить и теоретически со временем они или их потомки создали бы и развили свой язык – несмотря на то, что их никогда не учили ни одному языку. Более того, этот язык, хотя он был бы совершенно отличным от всех известных нам языков, поддавался бы анализу на базе тех же методов, которыми пользуются лингвисты для анализа других языков, и переводу на все известные языки. Но я бы развил этот аргумент еще дальше. Если бы наши новоиспеченные Адам и Ева могли выжить и иметь потомство – по-прежнему изолированные от влияния каких бы то ни было культур – то, в конце концов, они бы создали общество, в котором были бы: свои законы о собственности; правила, запрещающие инцест и регулирующие заключение браков; обычаи, регламентирующие различные табу и запреты; методы разрешения споров с минимальным кровопролитием; вера в сверхъестественное с соответствующими ритуальными действиями; система социального статуса и способов его проявления; церемонии посвящения молодых людей во взрослую жизнь; обычаи, принятые при ухаживании, включая применяемые женщинами способы украшения; системы символического украшения тела в общем; определенные мужские мероприятия и объединения, участие в которых женщин не допускается; различного рода азартные игры; промышленность, производящая оружие и орудия труда; мифы и легенды; танцы; внебрачные связи; а также, в различной дозировке – убийства, самоубийства, гомосексуализм, шизофрения, психозы и неврозы, а также различные доктора, извлекающие выгоду либо лечащие все это – в зависимости от того, с какой точки зрения на это посмотреть.
Культуру, в том числе наиболее блистательные и впечатляющие ее проявления в виде ритуальных и религиозных служб, можно интерпретировать как иерархическую систему приспособлений и устройств для отслеживания параметров среды. В главе 7 в качестве такой системы была описана совокупность биологических ответов – от сверхбыстрых биохимических реакций, протекающих за долю секунды, до замены одних аллелей генов другими, для чего требуется жизнь нескольких поколений. Тогда на этой схеме культура заняла место на “медленном” конце временной шкалы. Сейчас эту концепцию можно расширить. В той мере, в какой характерные особенности культуры являются негенетическими (ненаследственными), они могут быть отделены от биологической системы и расположены рядом с ней в качестве вспомогательной системы. Диапазон системы отслеживания исключительно по культурным параметрам почти полностью соответствует “медленному” сегменту системы отслеживания по биологическим параметрам и ранжируется от нескольких дней до поколений. К самым быстрым культурным откликам в промышленных цивилизациях относятся мода в одежде и стиле речи. Несколько более медленными являются политическая идеология и отношение общества к другим странам, а медленнее всего изменяются табу на кровосмесительные браки, а также вера и неверие в тех или иных небесных богов. Можно предположить, что культурные характеристики в большинстве своем адаптивны в дарвиновском смысле этого слова, даже если для каких-то из них этот процесс происходит опосредованно через выживание наиболее совершенно приспособленной социальной группы (Washburn and Howell, 1960; Masters, 1970). Второе предположение, которое стоит рассмотреть, чтобы завершить нашу биологическую аналогию, заключается в следующем: скорость изменения каждой конкретной совокупности поведенческих реакций в культуре отражает скорость изменения характеристик среды, с которыми связаны эти поведенческие реакции.
Формы культуры, которые изменяются медленно, имеют тенденцию к превращению в ритуал. Некоторые ученые-обществоведы провели аналогию между церемониями, принятыми в человеческом обществе, и тем, как в процессе коммуникации ведут себя животные. Это неверно. Большинство коммуникативных проявлений у животных является сигналами, значение которых весьма ограничено. Они соответствуют позам, выражениям лица, простейшим звукам человеческого параязыка. Лишь немногие проявления в поведении животных, такие, как самые сложные формы полового привлечения и перемены гнездования у птиц, являются настолько сложными и искусными, что подчас зоологи описывают их как “церемонии”. Но даже в этих случаях подобное сравнение неверно. В большинстве своем человеческие ритуалы имеют более глубокое значение, чем всего лишь непосредственная передача определенного сигнала. Как подчеркивал Дюркгейм (Durkheim), они не просто являются определенной меткой, но и заново подтверждают и обновляют моральные ценности данного общества.
Священные (духовные) ритуалы являются наиболее ярким примером ритуалов, принятых в человеческом обществе. В своих простейших формах они связаны с магией – активной попыткой манипулировать природой и богами. Росписи эпохи верхнего палеолита, найденные в пещерах Западной Европы, демонстрируют пристрастие к охоте на диких животных. На множестве рисунков изображены сцены охоты, с копьями и стрелами, вонзенными в тела жертв. На других рисунках изображаются люди, танцующие в шкурах и масках животных или стоящие перед животными со склоненными головами. Возможно, функциональным назначением этих рисунков было колдовство, основанное на внушении, логика которого заключается в том, что если сделать что-то с изображением, то же самое произойдет и с его прототипом в реальности. Такое упреждающее действие сравнимо с движениями демонстрации намерений у животных, которые в процессе эволюции зачастую ритуализировались, превращаясь в коммуникативные сигналы. “Виляющий” танец пчелы, напомним, является повторением в миниатюре того пути, который надо пролететь от гнезда до источника пищи. Примитивный человек, очевидно, с легкостью понимал, что означает это сложное поведение животных. Магией занимались, а в некоторых обществах и поныне занимаются особые люди, которых могли называть по-разному: шаманами, колдунами, знахарями. Считалось, что только они обладают тайным знанием и властью и могут иметь дело со сверхъестественными силами; эти люди зачастую обладали большим влиянием, чем вожди племени.
В формальной религии sensu stricto имеется много элементов магии, но она сосредотачивается на более глубоких, с более сильной родовой ориентацией верованиях. На уровне обрядов она проявляется в создании мифов, ритуалах умиротворения богов, в санкционировании принятых в обществе нравственных норм. Вместо шамана, управляющего физическими силами, существует священник, который сообщается с богами и молит их о милости в награду за почитание, жертвоприношения, а также надлежащее праведное поведение членов племени (сообщества). В более сложно устроенных сообществах государственное устройство и религия всегда естественным образом взаимосвязаны. Власть по праву помазанника божьего принадлежит королям, но высшее духовенство часто правит королями в силу того, что боги стоят выше королей.
Существует разумная гипотеза, согласно которой магия и тотемизм представляли собой результат непосредственной адаптации к среде и в общественной эволюции предшествовали религии как таковой. Сакральные традиции в человеческих обществах существуют практически повсеместно. Так же, как и мифы, объясняющие происхождение человека или, по крайней мере, то, какое отношение данный род имеет ко всему остальному миру. Но вера в небесных богов не является повсеместно распространенной. Всего у 28 из 81, то есть всего у 35 процентов сообществ, живших охотой и собирательством и изученных Уайтингом (Whiting, 1968), религиозные традиции предполагали веру в небесных богов. Концепция живого, нравственного бога, сотворившего мир, распространена еще меньше. Более того, эта концепция чаще всего свойственна тем обществам, которые ведут пастушеский образ жизни. Чем больше общество зависит от скотоводства, тем выше вероятность того, что для этого общества будет свойственна вера в бога-пастыря иудейско-христианского типа (см. табл. 26-3). В обществах других типов подобная вера встречается в 10 процентах случаев или еще реже. Кроме того, в монотеистических религиях Бог – всегда мужского пола. У этой сильно выраженной патриархальной тенденции есть несколько культурных предпосылок (Lenski and Lenski, 1970). Пастушеские общества в высшей степени мобильны, имеют четкую организацию и зачастую военизированы – то есть обладают чертами, изменяющими баланс власти в обществе в пользу мужчин. Важно также то, что скотоводство, составляющее основу экономики общества, является преимущественно мужской обязанностью. Так как иудеи были изначально пастушеским народом, в Библии Господь называется пастырем, а избранный народ – его овцами. Ислам – одна из самых строгих из существующих монотеистических вер – дал начало раннему государству, объединявшему скотоводческие народы Арабского полуострова. Тесная связь пастуха с его стадом, очевидно, создает микрокосм, в котором неизбежно возникают более глубокие размышления об отношении между человеком и силами, что стоят над ним.
Таблица 26-3. Религиозные верования в 66 земледельческих обществах, распределенных по группам в зависимости от доли скотоводства в их продовольственном обеспечении (по: Human Societies, by G. and Jean Lenski, 1970).
|
Доля скотоводства в продовольственном обеспечении, % |
Доля обществ, верующих в живого нравственного Бога-творца |
Количество обществ |
|
36–45 |
92 |
13 |
|
26–35 |
82 |
28 |
|
16–25 |
40 |
20 |
|
6–15 |
20 |
5 |
Хотя антропология становится все более и более сложной, она все же пока не дала повода ставить под сомнение заключение Макса Вебера о том, что более простые религии сводятся к поиску сверхъестественных сил, которые могут обеспечить получение совершенно мирских, земных благ – долгой жизни, достатка земли и пищи, избежания физических катастроф, а также поражения врагов. Формой группового выбора является и соперничество между различными религиозными сектами. Те из них, которые смогут привлечь последователей, выживут; те, что не смогут – нет. Следовательно, религии, как и другие институты человеческого общества, развиваются с целью содействия дальнейшему процветанию своих сторонников. Так как демографическое преимущество (преимущество в численности по сравнению с другими группами) получает вся группа в целом, это преимущество можно обеспечить отчасти посредством альтруизма и эксплуатации – средств, в результате применения которых определенные сегменты общества получат выгоду за счет других. И наоборот, демографическое преимущество может быть совокупностью в целом возросших индивидуальных способностей к адаптации (приспособленности). Возникшая в результате разница в социальных условиях будет разницей между более угнетающими и более благоприятствующими религиями. Возможно, все религии в определенной степени являются угнетающими, особенно если они пользуются поддержкой со стороны главы племени и различных сословий. Эту тенденцию еще более усиливает борьба и соперничество между различными обществами, так как религия с успехом можно приспособить к нуждам войны и экономического развития.
Неизменный парадокс религии состоит в том, что во многом суть ее представляется явным обманом, и все-таки религия остается движущей силой во всех обществах. Человек предпочитает верить, а не знать, и, как сказал Ницше, иметь в качестве цели пустоту, чем вообще не иметь цели. В конце века Дюркгейм опроверг представление о том, что такую силу можно на самом деле извлечь из “сотканных иллюзий”. И с тех самых пор ученые-обществоведы ищут свой психологический Розеттский камень, который должен дать ключ к раскрытию глубоко сокрытых тайных истин религиозного мышления. В своем проницательном аналитическом исследовании по данному вопросу Раппапорт (Rappaport, 1971) предположил, что буквально все формы духовных (сакральных) обычаев используются в коммуникативных целях. Кроме выполнения такой задачи, как институционализация моральных ценностей сообщества, эти церемонии могут давать информацию о силе и благосостоянии родов и семейств. У марингов (maring) Новой Гвинеи ни главы родов, ни какие-либо другие лидеры не могут приказать мужчинам племени доказать свою верность и принять участие в войне. Группа начинает исполнять ритуальный танец, и каждый из мужчин выражает свое желание оказать поддержку в военных действиях, вступая или не вступая в этот танец. Мощь военного консорциума можно определить, буквально пересчитав имеющихся воинов по головам. В более развитых сообществах той же цели служат военные парады, украшенные соответствующей атрибутикой и ритуалами государственной религии. Знаменитые церемонии индейцев Северо-Западного побережья позволяют членам общества продемонстрировать уровень своего благосостояния, раздавая определенное пропорциональное количество вещей бесплатно, в виде милостыни. Ритуалы также позволяют упорядочить взаимоотношения, в которых иначе была бы ненужная двусмысленность и неопределенность. Лучшими примерами этой модели коммуникаций являются rites de passage – церемонии посвящения. По мере взросления мальчика, его превращение из ребенка во взрослого происходит очень постепенно и в биологическом, и в психологическом смысле. Порой он будет вести себя как ребенок, хотя более адекватным с его стороны было бы взрослое поведение, и наоборот. Общество испытывает определенные проблемы в том, так или иначе следует его квалифицировать. Обряд посвящения устраняет эту неопределенность, произвольно изменяя классификацию и вместо непрерывного градиента вводя дихотомическое разделение. Кроме того, обряд служит для упрочения связей юноши с принимающей его группой взрослых.
Освятить, сделать священным какой-либо обычай или положение – значит, подтвердить, что они стоят вне сомнений, и установить наказание для любого, кто посмеет против них возражать. Духовное настолько отделено от мирского в повседневной жизни, что одно только повторение, воспроизведение его в неподобающей обстановке является нарушением, грехом. Такая крайняя степень санкционирования – суть любой религии – сопутствует тем традициям и догмам, которые обслуживают наиболее важные интересы данной группы. Сакральными ритуалами человека подготавливают к высшему служению и самопожертвованию. Под мощным воздействием особого языка, специальных костюмов, а также сакральных танцев и музыки, которые так точно воздействуют на эмоциональные центры человека, он получает “религиозный опыт”. Он готов доказывать свою преданность роду и семье, проявлять милосердие, посвящать себя духовному служению, отправляться на охоту, участвовать в сражении, умереть за Бога и страну. Deus vult – таким был воинственный клич участников первого крестового похода. Так повелел Бог, но весь основной выигрыш достался на долю суммарной дарвиновской приспособленности рода (племени) – именно она становится здесь основным “благоприобретателем”, пусть и незаметным.
Генри Бергсон (Henri Bergson) был первым, кто дал определение второй силы, обуславливающей формализацию морали и религии. Чрезвычайная пластичность социального поведения людей представляет одновременно и могущественную силу, и реальную опасность. Если бы каждая семья сама вырабатывала и устанавливала для себя правила поведения, результатом была бы невыносимая постоянная смена традиций и растущий хаос. Для противодействия эгоистичному поведению и “растворяющей способности” духовных и интеллектуальных лидеров, каждое сообщество должно иметь свой кодекс, систему значимых символов и ценностей. В широком смысле буквально любая совокупность соглашений лучше, чем вообще никакой. Так как действуют произвольно выбранные кодексы, организация имеет тенденцию быть неэффективной и искаженной излишней пристрастностью. Как это в краткой форме выражено у Раппапорта: “Освящение превращает произвольное в необходимое, и регулирующие механизмы, которые являются произвольными, скорее всего, окажутся освящены”. Процесс порождает критику, и в обществах с наиболее высокими уровнями грамотности и самосознания появляются мечтатели и революционеры, настроенные на изменение системы. Реформаторство сталкивается с подавлением и репрессиями, потому что в той степени, в какой правила являются освященными и мифологизированными, для большинства людей они находятся вне сомнений, а несогласие с ними определяется как богохульство.
Это приводит нас к вопросу, который в значительной степени биологический по сути своей: к вопросу об эволюции способности человека к индоктринации (Campbell, 1972). Человеческие существа поддаются индоктринации до смешного легко – они сами ищут этого. Если предположить в качестве аргумента, что способность к индоктринации может эволюционировать, то на каком уровне имеет место естественный отбор? Один крайний вариант заключается в том, что единицей отбора является группа. Когда ее соответствие остальному обществу становится слишком слабым, группа вымирает. При таком варианте эгоцентрически настроенные индивидуалисты–члены общества одерживают верх и быстро увеличивают свою численность за счет других. Но их растущее господство увеличивает уязвимость общества и ускоряет его вымирание. На место исчезнувших приходят общества с более высокой частотностью конформистских генов, увеличивая таким образом общую частоту встречаемости генов в метапопуляции обществ. Распространение генов будет происходить быстрее, если метапопуляция (например, комплекс племен) одновременно будет также увеличиваться в размере. Формальные модели этого процесса показывают, что если скорость вымирания общества достаточно высока по отношению к интенсивности нейтрализующего индивидуального отбора, то частота встречаемости генов альтруизма может увеличиться и стать умеренно высокой. Гены должны быть такие, чтобы они благотворно сказывались на способности к индоктринации, даже за счет членов общества–носителей этих генов. Например, готовность идти на смерть в бою может быть полезной для выживания группы благодаря тем генам, которые делают возможным проявление губительной военной дисциплины. Гипотеза группового отбора достаточно хорошо объясняет эволюцию восприимчивости к индоктринации.
В равной степени хорошо ее объясняет конкурирующая гипотеза отбора на индивидуальном уровне. Согласно этой гипотезе, способность каждого отдельного человека к конформизму позволяет всем людям вместе пользоваться благами и преимуществами членства в одном обществе с минимальным риском и оптимальными энергозатратами. Хотя их эгоцентричные конкуренты могут на какой-то момент добиться преимущества, в долгосрочной перспективе они проигрывают из-за своего остракизма и репрессивных проявлений. Конформисты совершают альтруистические акты, возможно, даже рискуют собственной жизнью, не из-за того, что на групповом уровне произошел отбор генов самоотречения, а потому, что время от времени группа может воспользоваться преимуществами восприимчивости к индоктринации, которая в других случаях благоприятна для личности.
Эти две гипотезы не являются взаимоисключающими. Групповой и индивидуальный отбор могут подкреплять друг друга. Если война требует спартанских качеств и приводит к гибели некоторых воинов, победа может с лихвой вознаградить тех, кто выживет, новыми землями, властью и возросшими возможностями продолжения рода. Среднестатистический человек выходит победителем в игре под названием “совокупная приспособленность”, получая выгоду потому, что совместными усилиями всех участников среднестатистическому члену общества достается больше благ, чем он сам мог бы вложить. (Более полное обсуждение социологии религии см.: E. O. Wilson, 1978).
Этика
Ученые и гуманитарии должны одновременно задуматься над тем, не пришла ли пора на время забрать этику из рук философов и “биологизировать” ее. В настоящее время предмет исчерпывается несколькими весьма разобщенными теоретическими построениями. Первое – этический интуитивизм, понятие о том, что разум обладает непосредственным знанием того, что истинно и что ложно, и что это знание разум может формализовать с помощью логики и перевести на язык законов и норм общественной деятельности. Чистейшей концепцией, которой долгое время руководствовалась гражданская мысль Запада, является теория общественного договора, сформулированная Локком, Руссо и Кантом. В наше время эта концепция обновлена и включена в качестве компонента цельной философской системы Джоном Ролзом (John Rawls, 1971). Его основная идея заключается в том, что справедливость и правосудие должны быть не просто частью системы государственного управления, но объектом первоначального, основного контракта. Принципы, которые Ролзом названы “справедливость как честность” (распределительная справедливость) – это принципы, на которых остановили бы свой выбор свободные и разумно мыслящие люди, если бы они образовывали общественное объединение на условиях равной выгоды и хотели бы определить фундаментальные правила своего союза. Оценивая правомерность принятых впоследствии законов и правил поведения, необходимо проверять их на соответствие тем самым изначально принятым неоспоримым фундаментальным правилам.
Ахиллесовой пятой в положениях интуитивизма является то, что он полагается на эмоциональные суждения, производимые головным мозгом, как будто обращаться с этим органом следует как с “черным ящиком”. Хотя мало кто будет возражать против того, что справедливость как честность по отношению ко всем – это идеальное состояние освобожденных от телесной оболочки, абстрактных душ, эта концепция не имеет никакой объясняющей и прогнозирующей силы в отношении мира человеческих существ. Она и не рассматривает основных последствий строгого соблюдения положений этой теории для среды или для генетического фонда. Возможно, в настоящем тысячелетии объяснения и предсказания не понадобятся. Но это маловероятно – генотип человека и экосистемы, в которых он обитает, отличаются огромным разнообразием характеристик. В любом случае, полное исследование механизмов высшей нервной деятельности, имеющих отношение к созданию этических суждений, необходимо и уже проводится. Примером является создание второй теоретической модели, которую можно назвать этическим бихевиоризмом. В основе ее лежит предположение, которое наиболее полно было высказано Скоттом (J. F. Scott, 1971): моральные обязательства полностью усваиваются в процессе обучения, причем основным механизмом является “выработка условных рефлексов” в процессе обучения методом проб и ошибок. Другими словами, дети просто усваивают поведенческие нормы, принятые в обществе. Этой теории противостоит эволюционно-генетическая теория нравственного поведения. Наиболее убедительно с помощью документальных доказательств ее подтвердил Лоуренс Колберг (Lawrence Kohlberg, 1969). Колберг – структуралист, находящийся, в частности, под явным влиянием Пиаже (женевской школы генетической психологии – прим. перев. А.В.), а потому он еще не был связан с остальной биологией. Для обозначения общей концепции Пиаже пользовался выражением “генетическая эпистемология”, а Колберг – “когнитивно-эволюционная”. Однако результаты исследований обоих в конце концов войдут в сферу расширенной эволюционной биологии и генетики. Метод Колберга заключался в том, чтобы записывать и классифицировать вербальные ответные реакции детей в процессе решения морально-этических задач. Он разграничил шесть последовательных стадий развития нравственно-этического мышления, через которые может проходить личность в процессе своего умственного и психического созревания. Ребенок начинает со стадии первоначально полной зависимости от внешнего контроля и управления и доходит до стадии все большего усложнения внутренне усвоенных норм и стандартов (см. табл. 26-4). До сих пор не был исследован вопрос о том, насколько пластичными и гибкими являются основополагающие законы и правила. Не оценена их межкультурная изменчивость а, следовательно, не определена их наследуемость. Различие между этическим бихевиоризмом и нынешней версией эволюционно-генетического анализа заключается в том, что первая теория постулирует механизм (выработка оперантного условного рефлекса) без каких бы то ни было доказательств, а последняя представляет доказательства, не оговаривая никаких механизмов. В основе этого несоответствия нет никакой концептуальной проблемы. Изучение нравственного развития – это всего лишь более усложненный и менее поддающийся обработке вариант проблемы генетической изменчивости (см. главу 2). При разрастании массива данных можно ожидать объединения этих двух подходов с формированием более или менее четкого метода из области генетики поведения.
Таблица 26-4. Классификация морально-нравственных суждений по уровням и стадиям развития (на основе данных Колберга; Kohlberg, 1969).
|
Уровень |
Основа морально-нравственного суждения |
Стадия развития |
|
I |
Нравственная ценностная установка определяется наказанием и поощрением |
1. Послушание, подчинение правилам и власти во избежание наказания |
| 2. Конформизм, соответствие нормам с целью получения вознаграждения и взаимовыгодного обмена |
||
|
II |
Нравственная ценностная установка состоит в том, чтобы исполнять правильную роль, соблюдать установленный порядок и оправдывать ожидания других людей |
3. Установка “ай да молодец”: конформизм с целью избежать недовольства и отрицания со стороны других |
| 4. Установка на исполнение обязанностей: конформизм с целью избежать осуждения со стороны властей, нарушения порядка и последующего обвинения |
||
|
III |
Нравственная ценностная установка заключается в том, чтобы соответствовать общим нормам и стандартам, соглашаясь с соответствующими правами и обязанностями |
5. Правовая установка: признание важности и ценности контрактов, определенный произвол в создании правил для обеспечения общего блага. |
| 6. Установка по совести или по принципам: верность принципам свободы выбора, которые могут иметь приоритет над законом, если соблюдение и применение закона приносит больше вреда, чем пользы. |
Даже если бы завтра решение проблемы было найдено, важная ее часть, тем не менее, оставалась бы неразрешенной. Это – генетическая эволюция морали. В первой главе этой книги я говорил о том, что философы, рассуждающие о проблемах морали и нравственности, интуитивно воспринимают этические каноны морали, полагаясь на реакции эмоциональных центров своей собственной гипоталамной системы. То же самое справедливо и в отношении эволюционистов, хотя сами они против этого категорически возражают. Смысл правил и канонов можно расшифровать, только если интерпретировать деятельность эмоциональных центров как проявление биологической адаптации. В некоторых аспектах эта деятельность, вероятно, устаревает, становясь пережитком приспосабливаемости к самой примитивной форме родовой организации. В каких-то аспектах она находится буквально в statu nascendi, образуя новые и быстро изменяющиеся формы адаптации к сельскохозяйственной и городской жизни. Возникающая в результате путаница усиливается другими факторами. В той мере, в какой односторонне альтруистичные гены сформировались в популяции в процессе группового отбора, противостоять им будут аллеломорфы, успешно прошедшие отбор на индивидуальном уровне. Конфликт импульсов, управляемых различными генными структурами, будет, вероятно, распространен в популяции широко, поскольку из современной теории следует, что гены сохраняются лучше всего в состоянии сбалансированного полиморфизма (глава 5). Амбивалентность морали будет в дальнейшем еще больше усилена тем обстоятельством, что существование определенной схемы, системы этических норм, связанных с половой и возрастной принадлежностью, может обеспечивать большую генетическую приспособленность, чем один-единственный кодекс морального поведения, применяемый одинаково ко всем половозрастным группам. Таким образом можно объяснить некоторые различия в стадиях, описанных Колбергом. Например, основанная на личных принципах самоцентристская позиция и относительное нежелание совершать альтруистические акты маленьких детей должны обеспечивать им определенное преимущество. Точно так же молодежь и подростки одного пола должны быть связаны более крепкими узами со своими ровесниками, а потому они, как правило, чувствительны к одобрению, выражаемому сверстниками. Причина в том, что в этот период наибольшее преимущество связано с формированием альянсов и повышением социального статуса – в отличие от более поздних периодов, когда половая и родительская мораль становится наиважнейшей детерминантой приспособленности. Генетически запрограммированные конфликты между представителями различных полов и между отцами и детьми по типу, описываемому моделями Трайверса (главы 15 и 16), также, вероятно, способствуют закреплению возрастных различий в моральных обязательствах различных видов и степеней. Наконец, моральные требования отдельных людей на ранних фазах колониального роста должны различаться по многим параметрам от требований людей, живущих в условиях демографического равновесия или в периоды перенаселения. Метапопуляции с высокими уровнями r-угасания будут иметь тенденцию к генетическому разнообразию и отличию от других видов популяции в своем этическом поведении (см. главу 5).
Если такая теория врожденного морального плюрализма хотя бы в какой-то мере соответствует истине, то необходимость эволюционного подхода к морали и этике самоочевидна. Должно быть также ясно, что ни одну отдельно взятую совокупность моральных требований и стандартов нельзя применять одинаково ко всем человеческим популяциям, за исключением классов, образованных по половозрастным признакам в рамках каждой популяции. Применить единый кодекс, таким образом, значит создать сложные, трудноразрешимые моральные дилеммы – такими, безусловно, является современное состояние человечества. (Более полное обсуждение применения социобиологии в сфере этики и антропологии опубл. в: Wilson, 1978; Stent, ed., 1978).
Эстетика
Творческие, художественные импульсы ни в коем случае не являются свойственными исключительно только человеку. В 1962 году, когда Десмонд Моррис рассматривал этот вопрос в своей работе “Биология искусства” (Desmond Morris, The Biology of Art), 32 обезьянам-приматам было предложено выполнить несколько рисунков и наскальных росписей. Среди обезьян были 23 шимпанзе, 2 гориллы, 3 орангутанга, 4 мартышки-капуцина. Ни одна из обезьян не подвергалась специальному обучению, единственное, что они все получили – это доступ к необходимым материалам и инструментам для рисования. На самом деле попытки руководить деятельностью животных, стимулируя их к подражательству, всегда заканчивались неудачей. Стремление использовать живописное и рисовальное оборудование было сильным, поэтому никакого поощрения от людей-наблюдателей обезьянам не требовалось. И молодые, и старые животные были так поглощены этим занятием, что предпочли его приему пищи и иногда отвечали вспышками раздражения или гнева, если их останавливали. Две шимпанзе, которых изучали особенно пристально, поработали в высшей степени продуктивно. “Альфа” сделала более 200 рисунков, а знаменитый “Конго”, который заслуживает звания Пикассо среди человекообразных обезьян, является автором почти 400. Хотя большинство творений представляли из себя каракули, делались они по шаблонам, которые никак нельзя было назвать случайными. Линии и пятна распределялись по всей поверхности чистого листа по направлению от фигуры, расположенной в центре рисунка, к внешнему краю. Если рисунок начинался на одной стороне чистой страницы, шимпанзе затем обычно переходила и на противоположную сторону листа, чтобы соблюсти равновесие. Со временем росчерки становились толще, начинаясь с прямых линий, они развивались в более сложные множественные штрихи. Образцы рисунков Конго становились все сложнее и сложнее, проходя те же стадии, что и рисунки очень маленьких детей, и следуя той же эволюционной схеме: переходя от линий к круговым диаграммам в форме лопастей вентилятора, и даже к полностью законченным кругам. Другие шимпанзе рисовали кресты.
Художественную деятельность шимпанзе вполне можно считать специфической реализацией навыков применения инструментов, свойственных этим обезьянам. Представители этого вида демонстрируют в общей сложности около 10 технических методик, каждая из которых требует навыков ручного труда. Возможно, все они были усовершенствованы в результате практического применения, в то же время, по крайней мере некоторые из них передавались как традиция от одного поколения к другому. Шимпанзе обладают достаточно высокими способностями в изобретении новых методов и технологий – таких, как использование палки для проталкивания предметов через прутья клетки и в качестве рычага для открывания коробок. Таким образом, тенденция манипулировать предметами и исследовать их возможное применение обеспечивают шимпанзе преимущество в процессе их адаптации.
Тем более это относится и к возникновению искусства у человека. Как показал Уошберн (Washburn, 1970), человеческие существа в течение 99% всего времени в своей истории занимались охотой и собирательством, и в течение этого времени каждому человеку приходилось изготавливать свои собственные орудия производства. Признание формы и качества исполнения было необходимым условием выживания, и это, возможно, обусловило также появление общественного одобрения. Обе формы успеха вознаграждались более высокой генетической приспособленностью. Если шимпанзе Конго удалось дорасти до стадии рисования простых диаграмм, не так уж трудно представить, как примитивный человек прогрессировал до уровня изображения фигур. Как только этот уровень был достигнут, переход к использованию искусства в магических обрядах и ритуалах, основанных на подражании, очевидно, последовал очень скоро. Искусство, очевидно, сыграло, таким образом, стимулирующую роль и в развитии культуры, и в развитии умственных способностей человека. В конце концов как идеографическое отображение языка возникло письмо.
Музыка в том или ином виде также производится некоторыми животными. Людям кажется прекрасным сложное певческое поведение птиц, которое служит целям ухаживания и обозначения территории – и, возможно, птичье пение нравится людям именно по тем причинам, по каким оно нужно самим птицам. Птицы могут ясно и четко определить по пению вид, физическое состояние и умственное развитие певца. Информационная наполненность и точность передачи настроения в неменьшей степени являются стандартами совершенного музыкального исполнения и у человека. Пение и танец служат для того, чтобы объединить группы, управлять эмоциями людей, подготовить их к совместным действиям. Карнавальные проявления у шимпанзе, описанные в предыдущих главах, в этом отношении очень напоминают человеческие празднества. Обезьяны бегают, скачут, стучат по стволам деревьев, выполняя движения, напоминающие движения барабанщиков, и громко кричат на всю округу. Эти действия, по крайней мере отчасти, служат для того, чтобы собрать группу, члены которой кормятся совместно. Эти действия могут напоминать церемонии самых древних человеческих племен. Тем не менее, в ходе последующей эволюции человеческого вида появились и коренные отличия. Людская музыка стала независимой от иконографии, точно так же, как и настоящий язык отделился от простейших ритуалов, характерных для общения в мире животных. Музыка обладает возможностями для безграничной и произвольной символизации, и в музыке используются те же правила построения фраз и их порядок, имеющий то же функциональное значение, что и синтаксис.
Защита территории и родовой строй
Антропологи часто не учитывают территориальное поведение как общую черту, свойственную человеку. Это происходит, когда из области зоологии заимствуется наиболее ограниченная концепция этого явления – поведенческая “модель колюшки”, для которой характерны встреча обитателей на фиксированных границах для осуществления угрозы и отпугивания друг друга. Но ранее в главе 12 я продемонстрировал, почему необходимо принять более широкое определение территории как любого ареала, занимаемого более или менее исключительно каким-либо животным или группой животных и охраняемой ими посредством прямой защиты или мечения. Методы отпугивания могут быть как четко выраженными, как прямая решительная атака, так и скрытыми, как помещение химического секрета желез в местах контрольных меток территории. В равной степени ответные реакции животных на действия соседей могут проявляться самыми различными способами. Для каждого вида характерна своя собственная особая поведенческая шкала. В своих крайних проявлениях эта шкала может включать реакции от открытой вражды, скажем, во время сезона размножения или в условиях увеличения плотности популяции, и вплоть до косвенных форм обозначения территории или даже до полного отсутствия территориальной поведенческой реакции как таковой. В менее экстремальных проявлениях шкала может варьировать только от умеренно агрессивного поведения при близком приближении соперника до призыва на более дальних расстояниях, или проявляться в различной степени интенсивности помещения меток, и так далее. Можно попытаться охарактеризовать поведенческую шкалу вида и определить параметры, которые заставляют животных двигаться вверх или вниз по этой шкале.
Если принять эти характеристики, есть основания сделать вывод о том, что территориальное поведение является общей чертой, характерной для сообществ, живущих охотой и собирательством. В интересном обзоре собранных им данных Эдвин Уилмсен (Edwin Wilmsen, 1973) обнаружил, что эти относительно примитивные сообщества в принципе не отличались в применяемых ими стратегиях землевладения от многих других видов млекопитающих. Регулярное проявление прямой агрессии характерны для меньшинства народов, живущих за счет охоты и собирательства – например, у племен чиппеуа (Chippewa), сиу (Sioux) и вашо (Washo) в Северной Америке и менгин (Murngin) и тиви (Tiwi) в Австралии. Достаточное пространство и демографическое равновесие обеспечивались за счет набегов, убийств, угрозы применения колдовских ритуалов. Племя вашо в Неваде активно защищало участки в ядре своих зон обитания, на территории которых они обустраивали свои зимние жилища. Менее ярко выраженные, косвенные формы взаимодействия могут давать такой же результат. Гун-бушмены из района Ньэй-Ньэй называют себя “совершенными” или “чистыми”, а других представителей племени гунов – “чужими” убийцами, применяющими смертельные яды.
Территориальное поведение человека иногда специализировано таким образом, который свидетельствует о его очевидном функциональном назначении. Не так давно, в 1930, бушмены области Доуб в юго-западной Африке ввели у себя закон эксклюзивного права семьи на владение земельным участком в период сезона дождей. Право распространялось только на сбор плодов; другим группам разрешалось в это время охотиться на том же участке на животных (R. B. Lee in Wilmsen, 1973). Другие племена и народы, добывающие свое пропитание охотой и собирательством, как оказалось, следовали тому же примеру и использовали двойной закон: более или менее эксклюзивное право родов или семей на пользование самыми плодоносными источниками (землями), и, наоборот, широкое совместное использование охотничьих территорий. Таким образом, исходное предположение Bartholomew and Birdsell (1953) о том, что австралопитек и примитивный хомо сапиенс были территориальными видами, по-прежнему является жизнеспособной гипотезой. Более того, по закону экологической эффективности, ареал и территории обитания были, возможно, велики, а плотность популяции, соответственно, низка. Этот закон, напомним, определяет, что если пищевой рацион состоит из животной пищи, то ареал, необходимый для добывания равного в энергетическом выражении пропитания, должен быть приблизительно в 10 раз больше, чем если пищевой рацион состоит из растительной пищи. Современные группы, добывающие пропитание охотой и собирательством и состоящие из приблизительно 25 человек, обычно занимают территорию в размере 1000–3000 кв. километров. Эта территория сравнима по размерам с участком обитания стаи волков, но не менее чем в 100 раз больше той, что занимает стая горилл, которые питаются исключительно растениями.
Исходя из предположения о территориальном поведении и защите территории, Ганс Каммер (Hans Kummer, 1971) сделал важное дополнение к пониманию поведения человека. Пространственные отношения между группами в природе просты и могут поддерживаться с помощью относительно небольшого числа простых методов проявления агрессии. Пространственные отношения и отношения доминирования внутри групп гораздо более сложны, они тесно связаны со всем остальным репертуаром общественных отношений. Отчасти проблема человека заключается в том, что межгрупповые ответные реакции все еще являются грубыми и примитивными и не соответствуют обширным и разнообразным взаимоотношениям вне территориальных границ, которых требует от человека жизнь в цивилизованном мире. Неутешительным результатом этого является то, что Гаррет Хардин (Garrett Hardin, 1972) определил как трайбализм, стремление к племенному обособлению в современном его смысле:
Любую группу людей, которая считает себя отдельной группой и которую внешний мир также воспринимает как отдельную группу, можно назвать племенем. Эта группа может быть отдельной расой в том понимании, какое обычно имеет это определение, но не обязательно; она точно так же может быть религиозной сектой, политической группой или профессиональной группой. Важной чертой племени является то, что оно имеет двойной морально-нравственный стандарт – один вид поведения принят для внутригрупповых отношений и другой – для внегрупповых отношений.
Одной из неблагоприятных и неизбежных черт трайбализма является то, что, в конце концов, он приводит к контр-трайбализму (или, другими словами, он поляризует общество, раскалывая его на два лагеря).
Опасаясь враждебных группировок вокруг себя, “племя” отказывается подчиняться общим интересам общества. Маловероятно, что оно добровольно согласится контролировать рост численности своей популяции. Как сингальцы и тамилы на Шри-Ланке, соперничающие племена могут даже вступить в гонку, соревнуясь, кто кого “победит” при размножении. Источники пропитания секвестированы. Такие ценности, как справедливость и свобода, приходят в упадок. Рост числа реальных и воображаемых врагов увеличивает чувство групповой идентичности и мобилизует членов племени. В политике значимую роль играет ксенофобия. Отношение к несогласным внутри самой группы становится более грубым и жестким. История изобилует примерами эскалации такого процесса до точки, когда в обществе происходит раскол или начинается война. Абсолютного иммунитета против этого нет ни в одной стране.
Ранняя социальная эволюция
Можно отметить в ментальной эволюции современного человека две явственные стадии ускорения. Первая произошла в период генезиса первых обезьянолюдей (австралопитеков) из крупных древесных форм приматов. Поскольку, как сейчас считается, примитивные гоминиды рамапитеки находятся на прямой линии наших предков, то эти изменения могли потребовать как максимум 10 миллионов лет. Австралопитеки жили 5 миллионов лет назад, и 3 миллиона лет назад разделились на несколько форм, включая, возможно, первых примитивных Homo (Tobias 1973). Как показано на рис 26-1, эволюция этих переходных гоминид сопровождалась ускорением роста объёма мозга. Одновременно сформировалось вертикальное положение тела, двуногая локомоция, а руки получили возможность выполнять точные хватательные движения. Нет сомнений, что эти ранние люди использовали орудия в намного большей степени, чем современные шимпанзе. Грубо обработанные каменные орудия, сделанные путём обкалывания и трения друг о друга уже появляются на местах стоянок.
Вторая, намного более энергичная фаза ускорения началась около 100 000 лет назад. Она включает в себя, прежде всего, культурную эволюцию, и, несомненно, вытекает из фенотипа, основанного на врождённом потенциале мозга, накопленном в предшествующие миллионы лет. Мозг достиг нового порога, за которым началась чрезвычайно быстрая умственная эволюция. Эта вторая фаза ни коим образом не была запланирована, просто это был момент раскрытия возросшего потенциала мозга.
Изучение происхождения человека можно соотнести с двумя вопросами, связанными с двумя стадиями умственной эволюции:
– Какие особенности окружающей обстановки побудили гоминид адаптироваться иначе, чем других приматов, и начать их уникальный путь эволюции?
– Раз начавшись, почему такая эволюция завела гоминид столь далеко?
Поиск первоначальных движущих сил ранней эволюции человека растянулся более чем на 25 лет. В число участников этого поиска входили: Dart (1949, 1956), Bartholomew и Birdsell (1953), Etkin (1954), Washburn и Avis (1958), Washburn et al. (1961), Rabb et al. (1967), Reynolds (1968), Schaller и Lowther (1969), C. J. Jolly (1970) и Kortlandt (1972). Эти авторы сконцентрировались на двух бесспорно важных фактах из биологии австралопитеков и ранних Homo. Первый состоит в том, что Australopithecus africanus, вид, который, скорее всего, был прямым предком Homo, жил в открытой саванне. Следы иноса от песчинок, обнаруженные на окаменелостях Стеркфонтейна (Sterkfontein) свидетельствуют о сухом климате, а находки свиней, антилоп и других млекопитающих ассоциированы с останками гоминид, что в таком случае точно указывают на существовании пастбищ. Образ жизни австралопитеков сформировался в условиях значительного изменения условий обитания. Их предки рамапитеки, как и их предшественники, жившие в лесах, были адаптированы к перемещению на деревьях посредством раскачивания на ветках. Лишь очень немногие другие приматы с относительно крупным телом были способны покинуть лес и проводить большую часть жизни на земле, на открытых местообитаниях. Нельзя сказать, что группы Australopithecus africanus большую часть времени бегали открыто. Какая-то их часть могла переносить свои игры в пещеры и даже жить в них как в постоянной резиденции, хотя убедительных указаний на то, что такое происходило часто, пока что нет (Kurtén, 1972). Какие-то группы могли перемещаться с наступлением ночи под защиту деревьев в небольших рощах, как это делают современные павианы. Важно указать, что большая часть, или даже все они находили пищу именно в саванне.
|
|
|
Рисунок 26-4. Упрощенный филогенез высших приматов Старого Света, показывающий, что только три существующих группы перешли от жизни в лесу к жизни в саванне. Это павианы (Papio), гелады (Theropithecus gelada) и люди. (Приводится по: Napier and Napier, 1967; Simons and Ettel, 1970). |
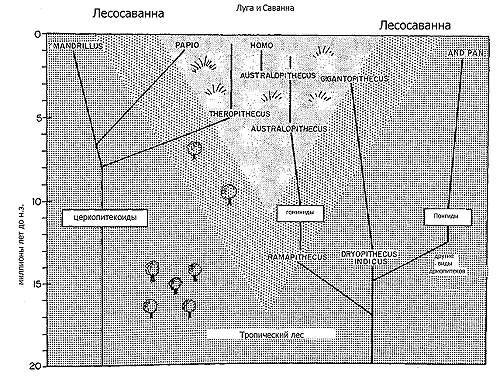
Вторая специфическая особенность экологии ранних людей состояла в степени их зависимости от животной пищи, несомненно большей, чем у любых ныне живущих обезьян. Австралопитеки употребляли самых разнообразных мелких животных. Места их стоянок содержат следы черепах, ящериц, змей, мышей, кроликов, дикобразов, и других мелких доступных им животных, в изобилии водящихся в саванне. Обезьянолюди также охотились на павианов с дубинками. Анализируя 58 черепов павианов, Дарт (Dart) обнаружил, что все они были умерщвлены ударом по голове, 50 – спереди, остальные – сзади. Австралопитеки также охотились на крупных животных, включая крупных сиватериев, жирафов, динотериев – хоботных с кривыми, спускающимися вниз бивнями, В ранне-Ашельскую эпоху, когда Homo erectus начал использовать каменные топоры, некоторые виды крупных африканских млекопитающих стали вымирать. Резонно предположить, что их несчастья произошли вследствие чрезмерной охоты на них все более опытными группами людей (Martin, 1966).
Какие выводы мы можем сделать из этих фактов о жизни ранних людей? Прежде чем попытаться дать ответ, мы должны отметить, что очень мало можно заключить непосредственно из сравнения с другими ныне живущими приматами. Гелады и павианы, живущие только в открытой местности формы – главным образом вегетарианцы. Они являют собой пример самое большее шести видов, которые слишком отделились один от другого в социальной организации, чтобы быть опорой для сравнения. Шимпанзе, более умные и социально изощренные обезьяны, живут в лесах, и тоже главным образом вегетарианцы. Их лишь эпизодические охотничьи предприятия могут показать нам поведение, могущее быть прямо соотнесенным с экологией и образом жизни, которые были значимы и для эволюции людей. Другие заслуживающие внимания особенности социальной организации шимпанзе, к примеру, быстроменяющийся состав подгрупп, межгрупповой обмен самками, замысловатый и продолжительный процесс общения, могут совпадать, а могут и не совпадать с таковыми у ранних людей. Мы не можем доказать, что эти особенности имеют экологические корреляции. В популярной литературе часто утверждается, что жизнь шимпанзе даёт ключ к пониманию жизни ранних людей. Это не обязательно верно. Сходные с человеческими особенности шимпанзе могут быть следствием эволюционной конвергенции, и в этом случае их привлечение для эволюционных реконструкций может ввести в заблуждение.
Наилучший способ, которому, как я полагаю, должны доверять большинство лиц, изучающих предмет, состоит в том, чтобы экстраполировать в прошлое жизнь охотничьих сообществ. Эта техника отражена в таблице 26-5. Используя синтез, отредактированный Lee и DeVore (1968; особенно смотри J. W. M. Whiting, страницы. 336–339), я перечислил наиболее общие черты охотничьих сообществ. Затем я оценил лабильность каждой поведенческой категории, указав число вариаций в категории, которая имеет место у видов приматов. Наименее лабильная категория – наиболее вероятная черта ныне живущих охотничьих сообществ, также наблюдалась и среди ранних людей.
Таблица 26-5. Особенности социальной структуры охотничьих сообществ и вероятность, что ими также обладали ранние люди.
|
Признаки, наблюдающиеся в жизни сообществ охотников и собирателей |
Вариабельность категории признака у других приматов |
Достоверность того, что ранние люди гомологично обладали теми же признаками |
|
Размер локальной группы: |
||
|
Не более 100 |
Очень вариабельна, но в пределах 3–100 |
Очень возможно 100 или меньше, но не очень достоверно |
|
Семья как ядро группы |
Очень вариабельно |
Не достоверно |
|
Разделение труда между полами |
||
|
Женщина – собиратель, мужчина – охотник |
Среди ныне живущих приматов – только у людей |
Не достоверно |
|
Самцы доминируют над самками |
Широко распространено, но не всеобще |
Достоверно |
|
Долговременные половые связи (брак); почти универсально; главным образом – полигиния |
Очень вариабельно |
Не достоверно |
|
Общая экзогамия, поощряемая брачными обычаями |
Среди ныне живущих приматов – только у людей |
Не достоверно |
|
Частые смены состава подгрупп (принцип расщепления–синтеза) |
Очень вариабельно |
Не достоверно |
|
Существенная территориальность, особенно в высокопродуктивных ареалах |
Наблюдается широко, но вариабельна в конкретных проявлениях |
Возможно, имела место. Конкретные проявления неизвестны |
|
Игровое поведение, особенно тренирующее сноровку, но не стратегию |
Практикуется повсеместно, по крайней мере – в простейших формах |
Очень достоверно |
|
Длительная материальная поддержка, выраженная социализация молодежи, расширенные отношения между матерью и детьми, особенно - между матерью и дочерьми |
Наблюдается повсеместно среди высших обезьян Старого Света |
Очень достоверно |
Что мы можем заключить с определённой степенью достоверности, так это то, что ранние люди жили маленькими территориальными группами, внутри которых самцы доминировали над самками. Интенсивность агрессивного поведения и природа его остаются неизвестными. Материальная поддержка была длительной, и отношения были до какой-то степени семейственными. Размышления об остальных аспектах социальной жизни не подкреплены анализом данных по лабильности и следовательно более шатки. Похоже, что ранние гоминиды добывали пищу группами. Судя по поведению павианов и гелад, такое поведение нужно соотнести с проблемой защиты от крупных хищников. В то же время австралопитеки и ранние Homo начали питаться крупными млекопитающими, групповая охота почти наверняка выгоднее, и даже необходима, подобно африканским диким псовым. Но это не есть неотразимый довод для заключения, что пока мужчины охотились, женщины сидели дома. Так делается сегодня в охотничьих сообществах, но сопоставление с другими приматами предлагает ключ, как и когда эта особенность проявлялась. Разумеется, нет существенных доводов предполагать априори, что самцы должны быть специализированным классом охотников. Самцы шимпанзе практикуют охоту, и это наводит на мысли. Но в отношении львов эти мысли нужно передумать, ибо львицы тоже добытчики, часто действующие в группе, и часто с детёнышами “на хвосте”, в то время как самцы держатся сзади. У африканских диких псовых оба пола равноправны в охоте. Это не говорит о том, что самцовая групповая охота не была ранней особенностью гоминид, но только то, что нет сильных независимых свидетельств в пользу этой гипотезы.
Всё это приводит нас к преобладающей теории об источниках социальности человека. Она содержит набор взаимосвязанных моделей, основанных отчасти на археологических свидетельствах, отчасти – на экстраполяциях в прошлое существующих охотничьих сообществ, и сравнении с другими ныне живущими видами приматов. Ядро этой теории можно приблизительно назвать “автокаталитической моделью”. Она предполагает, что когда древнейшие гоминиды перешли к двуногому передвижению, как составной части их адаптации к жизни на поверхности земли, их передние конечности освободились, что облегчило изготовление орудий и манипуляции с предметами, а разумность возрастала как составная часть навыков использования орудий. Умственные способности и привычка использовать инструменты взаимно усиливались, развивая материальную культуру. Кооперация в ходе охоты достигла совершенства, дав новые стимулы к эволюции умственных способностей, что в свою очередь позволило использовать более изощрённые инструменты, и так далее по нарастающей. В какой-то момент, возможно во времена поздних австралопитеков, или во время перехода австралопитеков в Homo, этот автокатализ подвёл развивающиеся популяции к определённому порогу развития, за которым гоминиды стали способны добывать антилоп, слонов, и других крупных травоядных млекопитающих, которыми в изобилии кишели африканские равнины. Вполне возможно, что процесс начался тогда, когда гоминиды учились отгонять крупных кошачьих, гиен, и других плотоядных от их добычи. (см рис 26-5). Тогда же они стали сами преимущественно охотниками, и были вынуждены защищать свою добычу от других хищников и падальщиков. Автокаталитическая модель обычно включает теорему о том, что переход охоте на крупную дичь ускорил процесс развития мозга. Этот переход даже мог быть той движущей силой, что обеспечила переход от австралопитековых предков к ранним Homo около двух миллионов лет назад. Другое предположение гласит, что самцы стали специализированными охотниками. Забота о детях облегчалась тесным социальным взаимодействием между самцами, которые уходили из постоянных жилищ на охоту, и самками, которые следили за детьми, и обеспечивали основное пропитание растительной пищей. Многие существенные детали полового поведения и семейной жизни легко определяются из этого базового разделения труда. Но эти детали не существенны для автокаталитической модели. Они добавлены, так как они наблюдаются в современных охотничьих сообществах.
|
|
|
Рисунок 26-5. На пороге автокаталитической социальной эволюции два миллиона лет назад, группы первых людей (Homo habilis) собирают пищу в африканской саванне. На этой умозрительной реконструкции группа изображена в процессе отгона конкурирующих хищников от только что павшего динотерия. Крупное слоноподобное существо пало от изнеможения или болезни, его кончина, возможно, была ускорена атакой животных, изображенных неподалёку. Люди только что появились на месте события. Некоторые отгоняют прочь хищников различными криками, размахиванием руками, палками, бросанием камней, в то время как отдалённые особи слева приготовились вступить в драку. Справа самка саблезубой кошки (Homotherium) и два ее подросшие детеныша испугано отпрянули (по крайней мере – временно). На их мордах угрожающе широко раскрыты пасти. Слева на переднем плане группа пятнистых гиен (Crocuta) также ретируются, но готовы ринуться обратно в первый же удобный момент. Люди весьма маленькие, меньше 1.5 метров в высоту, и по отдельности – не соперники крупным хищникам. Согласно превалирующей теории, высокая степень кооперации была, следовательно, необходима, чтобы завладеть такой добычей, и она развилась в тесной связи с высокими умственными способностями и превосходной способностью использовать орудия. На заднем плане можно видеть обстановку района Олдувай в нынешней Танзании, как она могла выглядеть в то время. Местность была холмистой, и окаймлённой с востока вулканическим высокогорьем. Популяции травоядных были плотными и разнообразными, как и сейчас. Слева на заднем плане видны трехпалые лошади (Hipparion), а справа – стадо гну и гигантских жирафоподобных существ, сиватериев. (Рисунок Сары Лэндри (Sarah Landry); с консультацией F. dark Howell. Реконструкция Homotherium основана частично на фигурке культуры Ориньяка; см.: Rousseau, 1971). |

Несмотря на внутреннюю непротиворечивость, автокаталитическая модель содержит возбуждающий любопытство пробел – что послужило пусковым механизмом? Самоподдержание уже начавшегося процесса легко объяснимо, но что его запустило? Почему древние гоминиды перешли к двуногости, вместо того, чтобы продолжать бегать на всех четырёх, подобно павианам или геладам? Клиффорд Джолли (Clifford Jolly, 970) предложил в качестве первоначального толчка специализацию на семенах трав. Из-за того, что самые ранние пра-люди, возможно ещё типа рамапитеков, были крупнейшими приматами, питавшимися семенами, преимущество у них получала способность манипулировать очень маленькими в сравнении с размерами ладони объектами. Короче, люди стали двуногими, чтобы сподручнее было срывать семена. Эта гипотеза – не беспочвенная фантазия. Джолли указывает на некоторые конвергентные особенности черепа и строения зубов сходные у людей и гелад, которые питаются семенами, насекомыми и другими мелкими объектами. Более того, гелады уникальны среди обезьян Старого Света и человекообразных тем, что подобно человеку имеют следующие эпигамные анатомические особенности: растущие волосы вокруг лица и шеи у самцов, и бросающиеся в глаза мясистые украшения на груди самок. Согласно модели Джолли, освобождение рук у ранних гоминид было преадаптацией, позволившей расширить употребление орудий и попутно катализовало эволюцию мозга и хищное поведение.
Позднейшая социальная эволюция
Автокаталитические реакции в реальных системах никогда не усиливаются беспредельно. Биологические параметры в норме меняют параметры среды так, чтобы замедлить эту реакцию или даже остановить ее. И почти сверхъестественно, что этого до сих пор не случилось в эволюции человека! Увеличение объёма мозга и совершенствование каменных орудий свидетельствуют о постепенном развитии умственных способностей в течение плейстоцена. С появлением Мустьерской культуры у неандертальцев примерно 75 000 лет назад, тенденции усилились, приведя в Европе к появлению верхнепалеолитической культуры у человека современного вида около 40 000 лет назад. Появившись около 10 000 лет назад, сельское хозяйство было усовершенствовано и широко распространено, население чрезвычайно увеличило свою плотность, и примитивные группы охотников встали на путь неумолимого роста племён, княжеств и государств. В итоге, после 1400 года цивилизации европейского типа изменили движущие силы прогресса, а знания и технологии растут не экспоненциально, а даже сверхэкспоненциально (смотри рис. 26-6, 26-7).
|
|
|
Рисунок 26-6. Четыре принципиальных типа сообществ в возрастающем порядке социополитической сложности, с существующими и исчезнувшими примерами в каждом. Некоторые социополитические институции из показанных, в приблизительном порядке расположены так, как они появлялись. (Заимствовано из: Flannery, 1972). |
|
|
|
Рисунок 26-7. Количество важных изобретений и открытий по столетиям, с 1000 до 1900 годы. (Заимствовано из: Lenski and Lenski, 1970). |
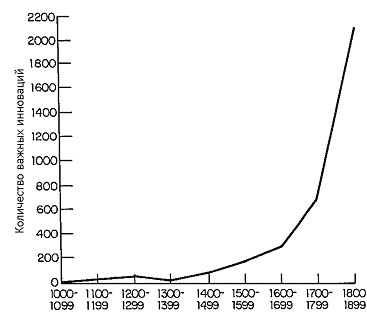
Нет оснований полагать, что в течение этого финального спринта случится остановка развития мыслительных способностей или появление склонности к особому социальному поведению. Теория популяционной генетики и эксперименты над другими организмами показывают, что существенные изменения могут происходить в течение менее чем 100 поколений, что в масштабах человеческой жизни примерно равно времени существования Римской империи. Две тысячи поколений приблизительно сменилось с момента появления человека современного типа в Европе; этого времени достаточно для появления новых видов, формирования их основных черт. Хотя мы не знаем, как развитие умственных способностей происходило в деталях, но вряд ли стоит полагать, что современные цивилизации всецело построены на капиталах, накопленных в течение уныло долгого плейстоцена.
Поскольку генетические и культурные следящие системы оперируют параллельными каналами, мы можем обойти их уникальность в какой-то момент, и вернуться к вопросу о первоначальных двигателях в поздней социальной эволюции человека в самом широком её смысле. Питание семенами – вполне состоятельное объяснение причин перемещения гоминидов в саванну, переход к охоте на крупную добычу может полагаться причиной эволюции в стадию Homo erectus. Но могла ли адаптация к групповой охоте в достаточной мере провести эволюцию гоминид по всему пути до Homo sapiens и далее, к сельскому хозяйству и цивилизации? Антропологи и биологи не считают, что этого достаточно. Они предлагают ряд других факторов, которые могут действовать как по отдельности, так и в совокупности.
ПОЛОВОЙ ОТБОР
Фокс (Fox, 1972), вслед за Чансом (Chance, 1962), выдвинул версию, что половой отбор мог быть дополнительным мотором, двигающим эволюцию человека на всём пути к человеку разумному. Его доводы вытекают из следующего: Полигиния – это общий признак живущих охотой групп, и могла также быть верной для сообществ ранних гоминид. Если это так, то преимущества могли быть получены при половом отборе, включающем как эпигамную демонстрацию, адресованную самкам, так и соперничество между самцами. Отбор мог быть улучшен демонстрацией постоянной готовности к спариванию, которая появилась у самок в связи с продолжительной сексуальной восприимчивостью. Вследствие существования высокого уровня кооперации внутри группы, унаследованной как изначальная адаптация австралопитеков, половой отбор мог быть нацелен на поощрение охотничьей доблести, лидерских качеств, навыков изготовления орудий и других видимых признаков, содействующими успеху семьи и всей группы, в которую входит некий самец. Агрессивность ограничивалась, и древние формы открытого доминирования приматов заменялись сложными социальными навыками. Молодые самцы находили выгодным соответствовать требованиям группы, управляя своей сексуальностью и агрессивностью, в расчёте, что это поможет им в лидерских устремлениях. В результате, доминирующий самец в сообществе гоминид, скорее всего, обладал мозаикой качеств, отражающих потребности в компромиссах. Как это чётко выразил Роберт Фокс, самец имел “самообладание, хитрость, организаторские способности, привлекательность для женщин, доброту к детям, снисходительность, упорство, красноречие, всё знал и умел в области самообороны и охоты”. Поскольку между этими изощрёнными социальными качествами и репродуктивным успехом возникает положительная обратная связь, социальная эволюция может неограниченно продолжаться без дополнительных селективных давлений со стороны внешней среды.
УМНОЖАЮЩИЕ ЭФФЕКТЫ КУЛЬТУРНЫХ НОВШЕСТВ И РАСШИРЕНИЯ ВОЗМОЖНОСТЕЙ ОБЩЕНИЯ
Что бы ни было основным двигателем, эволюция культуры осуществлялась за счёт роста возможности и готовности к обучению. Расширение возможностей контактов между особями и группами могли также возрастать. Можно предположить, что достижение критической массы культурных возможностей и сферы общения, в которых это стало выгодно для групп, активно расширяла и то и другое. Другими словами, обратная связь стала положительной. Этот механизм, подобно половому отбору, не требует дополнительных исходных данных сверх предела самого социального поведения. Но в отличие от полового отбора, он, возможно, достиг предельного уровня автокатализа очень поздно в предыстории человечества.
УВЕЛИЧЕНИЕ ПЛОТНОСТИ НАСЕЛЕНИЯ И СЕЛЬСКОХОЗЯЙСТВЕННОЕ ПРОИЗВОДСТВО
Общепринятый взгляд на развитие цивилизации обычно полагает, что инновации в области сельского хозяйства приводят к росту популяции, спокойному досугу, появлению класса незанятых людей и изобретателей, меньшую напряжённость борьбы за существование. Эта гипотеза существенно поблекла после открытия, что в племенах Кунг и других охотничьих сообществах люди работают меньше, и большее время проводят в досужих развлечениях, чем большинство фермеров. Примитивные сельскохозяйственные народы в общем случае не производят излишков, если к этому их не принуждают политические или религиозные авторитеты (Carneiro, 1970). Boserup (1965) пошел ещё дальше и предположил обратную взаимозависимость – именно рост плотности популяции вынуждает сообщества расширять и совершенствовать агрокультуру, а не наоборот. Однако это не объясняет первоначального роста плотности популяции. Охотничьи сообщества пребывают в примерном демографическом равновесии сотни тысяч лет. Что-то другое подтолкнуло их к тому, чтобы стать первыми фермерами. Вполне возможно, что решающим событием было всего лишь достижение некоего уровня интеллекта и счастливая случайная встреча с дикорастущими съедобными растениями. Раз запустившись, сельскохозяйственная экономика позволила увеличить плотность населения, что, в свою очередь, расширило сферы социальных контактов, технологический прогресс, и дальнейшую зависимость от сельского хозяйства. Немногие инновации, такие как изобретение колеса и ирригации, интенсифицировали этот процесс до той точки, от которой уже не было возврата.
ВОЙНЫ
На протяжении всей писанной истории ведение войны между племенами, тираниями и государствами было всеобщей практикой . Когда Сорокин (Sorokin) анализировал историю 11 европейских стран в течение от 275 до 1025 лет, он обнаружил, что в среднем они были вовлечены в различные виды военных акций в течение 47 процентов всего времени, или каждый год из двух. Значения различались от 28 процентов в случае Германии, до 67 процентов в случае Испании. Ранние тирании и государства Европы и среднего востока быстро сменяли друг друга, причём большинство этих столкновений носили характер геноцида. Экспансия генов всегда имела высшую важность. К примеру, после завоевания Мидианитян, Моисей повелел поступить, по сути, так же, как поступают самцы обезьян-лангуров, демонстрирующие агрессию и генетическую узурпацию:
Итак убейте всех детей мужеского пола, и всех женщин, познавших мужа на мужеском ложе, убейте; а всех детей женского пола, которые не познали мужеского ложа, оставьте в живых для себя. (Числ. 31, 17–18).
И веками позже, фон Клаузевиц призывал своего ученика, принца Прусской короны, по смыслу к тому же, к познанию биологической радости войны:
Будь дерзким и хитрыми в своих планах, решительным и стойким в своих действиях, нацеленных на славный конец, и удача вознаградит твой молодой лоб сверкающей славой, окружит принцессами, и запечатлеет твой образ в сердцах твоих будущих потомков.
Возможность того, что эндемичная война и генетическая узурпация могла быть эффективной силой в групповом отборе, была чётко осознана Чарльзом Дарвином. В “Происхождении человека” он предложил выдающуюся модель, предвосхитившую многие элементы современной теории группового отбора:
Итак, если некий мужчина племени умнее других, и изобрел новую ловушку или оружие, или иные средства нападения или защиты, даже в своих собственных интересах, то, не прибегая к принудительному убеждению, мог показать другим членам образец для подражания, которым все могли, следовательно, пользоваться. Вхождение в привычную практику каждого нового навыка в чём-то подобно усилению интеллекта. Если изобретение было важным, это племя могло увеличить свою численность, расширить свой ареал и вытеснить другие племена. И как дальнейшее следствие, племя могло ещё увеличить свою численность и повысить шансы того, что в нём родятся столь же выдающиеся и изобретательные члены. Если такой мужчина передаст своим детям своё умственное превосходство, то шансы на рождение столь же изобретательных членов могли бы быть несколько лучше, а в очень маленьком племени – несомненно лучше. Даже если лично у него не будет детей, то племя всё равно имело бы в своём генотипе эти качества, в чём можно удостовериться на примере скотоводов, которые защитой и разведением семей животных, которые затем забиваются, добились появления у них желаемых ценных качеств.
Дарвин видел, что групповой отбор мог не только усиливать отбор индивидуумов, но мог и противодействовать ему, что временами преобладало, особенно если размер размножающейся группы был невелик, и, следовательно, все в этой группе были недалёкие родственники. В сущности, подобная тема была позже глубоко раскрыта такими авторами как Keith (1949), Bigelow (1969), and Alexander (1971). Эти авторы представляли себе некоторые “благородные” черты человечества, такие как взаимовыручка, альтруизм, патриотизм, мужество на поле боя, и так далее, как генетическое следствие практики ведения войн.
Добавляя дополнительный постулат порогового эффекта, мы можем объяснить, почему процесс этот действовал лишь исключительно в эволюции человека (Wilson, 1972a). Если некое социальное хищное млекопитающее достигнет конкретного уровня интеллекта (а ранние гоминиды, будучи крупными приматами, было особенно предрасположены к этому), то некая группа имела бы возможность сознательно оценивать важность примыкающих социальных групп и иметь с ними дела упорядоченно-интеллигентным образом. Группа могла далее повлиять на соседнюю группу, присоединить ее территорию и увеличить собственную генетическую представительность в метапопуляции, сохранить в памяти племени этот успешный эпизод, повторяя его, расширяя географические пределы его встречаемости и быстро расширить свое влияние далее в метапопуляции. Так примитивные культурные возможности позволили бы распространиться конкретным генам. В ответ культурные возможности могли расширить распространение генов в генотипе метапопуляции. Раз начавшись, такая взаимная поддержка могла быть необратимой. Лишь комбинации генов способны дать превосходящее соответствие в соревновании с агрессивным геноцидом и могли также быть продуктом более эффективной техники агрессии, или иных возможностей реализации геноцида путём тех или иных форм мирных маневров. Как следствие, также возможен умственный и культурный прогресс. Вдобавок к автокаталитичности, такое развитие имеет интересную особенность – необходим отбор эпизодов лишь очень редких, чтобы продолжить отбор индивидуумов. По данной теории, геноцид и геносорбция сильно зависят от частоты появления агрессоров, которые должны появляться лишь изредка в немногих поколениях, чтобы направлять эволюцию. Эти немногие способны поддержать высокие частоты генов истинного альтруизма в группе (смотри главу 5). Изменчивость карты племен и княжеств, оцененная по картам истории ранней Европы и среднего востока (к примеру, атлас McEvedy, 1967 ) показывает достаточную величину дифференциации состояния групп для достижения данного эффекта. Более того, следует ожидать, что некоторые изолированные культуры будут избегать этого процесса для отдельных поколений, в сущности, временно возвращаясь к состоянию, которое этнографы классифицируют как состояние мира.
МНОГОФАКТОРНЫЕ СИСТЕМЫ
Каждый из вышеупомянутых механизмов по отдельности предположительно мог бы быть достаточным фактором социальной эволюции. Но их намного больше одного, и все действуют совместно, с различной силой и в сложном взаимодействии. Следовательно, наиболее реалистичная модель может быть вполне кибернетической, с причинами и следствиями, которые меняются местами в субциклах, в большой степени перекрывающихся. Одна такая схема, предложенная Adams (1966) для появления государств и городских сообществ, представлена на рис. 26-8. Излишне говорить, что уравнения для перевода в эту и подобные модели не написаны, и величины коэффициентов нельзя даже предположить для современной эпохи.
|
|
|
Рисунок 26-8. Мультифакторная модель происхождения государств и городских сообществ. (Заимствовано из: Flannery, 1972 на основе: Adams, 1966). |
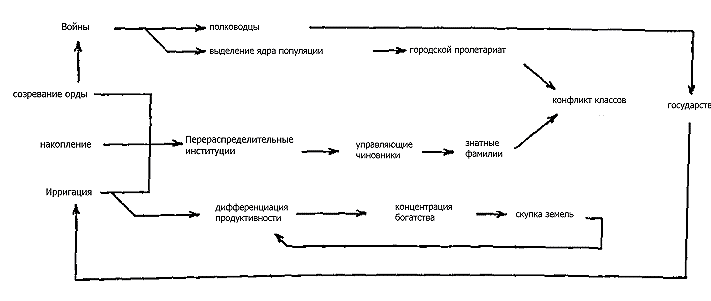
Как в однофакторных, так и в мультифакторных моделях социальной эволюции постулируется рост силы действия внутренних факторов. Этот сдвиг рассматривается как базис двухстадийного ускорения, упомянутого ранее. В начале эволюции гоминид, первоначальным двигателем эволюции было давление факторов внешней среды, такое же, какое двигает социальную эволюцию других животных видов. В некий момент, что представляется разумным предположением, гоминиды претерпели две фазы адаптации в своём развитии: первая вызвана переходом к жизни на открытых пространствах и к питанию семенами, и вторая, являющаяся преадаптацией анатомических и психических перемен, связанных с питанием семенами, вызвана переходом к охоте на крупных млекопитающих. Охота на крупную дичь вызывала дальнейший рост умственных способностей и совершенствование социальной организации, что подвело гоминид к порогу автокатализа – фазе эволюции, зависящей уже от внутренних причин. Эта вторая фаза – именно та, в которой появились наиболее выдающиеся особенности человека. Однако из давления этих особенностей я не хотел бы выводить последующую независимость социальной эволюции от факторов внешней среды. Железные законы демографии всё ещё сдерживают расширение популяции гоминид, и наиболее захватывающие культурные достижения появлялись как изобретение новых способов управления окружающей средой. Случилось так, что ментальные и социальные перемены пришли к большей зависимости от внутренней реорганизации и к меньшей – от прямых воздействий окружающей среды. Короче – социальная эволюция получила свой собственный мотор.
Будущее
Когда человечество достигнет экологически стабильного состояния, возможно, к концу 21-го века, интернализация социальной эволюции будет почти закончена. К этому времени биология будет на пике популярности, с быстро созревшими социальними науками. Некоторые историки науки напишут книги в этом свете, доказывая, что ускорение темпа открытий в этих областях предполагает более быстрые исследования. Но исторические прецеденты ввели нас в заблуждение ранее: тема, которую мы сейчас обсуждаем труднее физики и химии как минимум на два порядка величины.
Рассмотрим перспективы социологии. Эта наука сейчас находится в стадии естественной истории в своих исследованиях. Были попытки построить систему, но, как и в психологии, они оказались поспешными и дали мало что. Многие из этих шагов в теоретической социологии сегодня фактически лишь описывают явления и понятия, на манер естественной истории. Процесс анализа труден, ибо фундаментальные блоки трудноуловимы и возможно не существуют. Синтез в общем случае содержит скучный поиск по различным множествам дефиниций и метафор, нагромождённых более впечатлительными мыслителями (см. например: Inkeles, 1964; Friedrichs, 1970). А это тоже типичная фаза естественной истории.
С ростом возможностей описаний и экспериментов, социология движется с каждым днём к культурной антропологии, социальной психологии и экономике, и скоро поглотится ими. Эти дисциплины – фундамент для социологии sensu lato и очень вероятно, сформулируют первые феноменологические законы. Фактически, многие жизнеспособные качественные законы уже существуют. Они включают проверяемые утверждения о следующих взаимоотношениях: эффекты враждебности и стресса на этноцентризм и ксенофобию (LeVine and Campbell, 1972); положительная меж- и внутрикультурная корреляция между войной и боевыми видами спорта, приводящая к исключению гидравлической модели агрессивных действий (Sipes, 1973); точные, но пока специализированные модели продвижения и возможностей в профессиональных группах (White, 1970) и, далеко не последние по значимости, общие модели экономики.
Переход от чисто феноменологической к фундаментальной теории в социологии должен предполагать полное нейронное описание мозга человека. Только когда машину можно будет раскидать на бумаге до уровня клеток и снова собрать, тогда прояснятся свойства эмоций и нравственных суждений. Для оценки пределов поведенческих реакций и точности их гомеостатических механизмов могут быть использованы имитационные модели. Стресс будет быть описан в терминах нейрофизиологических возбуждений и торможений. Когнитивные способности будут переведены в графики. Обучение и творчество будут определены как изменения определенных частей когнитивной машины, управляемые входящими сигналами от эмоциональных центров. Поглотив психологию, новая нейробиология будет источником прочного набора главнейших принципов для социологии.
Роль эволюционной социобиологии в этом предприятии двояка. Она будет пытаться реконструировать историю нашей машины и определять адаптивную значимость каждой из его функций. Некоторые из этих функций почти наверняка устарели, будучи созданы для условий, актуальных в плейстоцене, таких как охота и межплеменные войны. Другие могут оказаться и ныне адаптивными на уровне особей и семей, но неадаптивными на уровне групп, или наоборот. Если принято такое решение, и культура формируется в соответствии с требованиями неизменной экологической обстановки, некоторые особенности поведения могут изменяться под влиянием жизненного опыта без вреда эмоциональной сфере или творческому потенциалу. Другие же – нет. Неопределенность в этом вопросе означает, что скиннеровским мечтаниям о культуре, предопределённой свыше для счастья, придется дождаться появления новой нейробиологии. С генетически точным и, следовательно, полностью ясным “кодом” этики тоже придется подождать.
Второй стороной вклада эволюционной социобиологии будет наблюдение за генетической основой социального поведения. Оптимальные социоэкономические системы никогда не могут быть совершенны – как следствие “теоремы невозможности Эрроу”, а также, видимо, из-за множественности врождённых этических стандартов. Более того, можно ожидать, что генетическая база, на которой построены такие нормативные системы, будет постоянно меняться. Человечество никогда не останавливается в своём развитии, в смысле, что в его популяциях происходит генетический дрейф. Спустя несколько поколений, эффект такого дрейфа может изменить идентичность социоэкономического оптимума. В частности, скорость потока генов по всему миру возросла до драматического уровня, и всё ускоряется, и значение коэффициентов родства внутри локальных сообществ соответственно уменьшается. Результатом может быть возможное сокращение альтуристического поведения вследствие утраты адаптивного смысла и потери генов, прежде закрепляемых групповым отбором (Haldane, 1932; Eshel, 1972). Как было показано ранее, когда поведенческие особенности подавлены или когда их изначальная функция стала нейтральной в адаптивном смысле, они подвергаются отрицательному отбору по принципу метаболитической консервации. Такие признаки могут в дальнейшем исчезнуть из популяции уже за десять поколений, или всего за два или три века применительно к человеку. При нынешнем недостаточном понимании человеческого мозга, мы не ведаем, какие из наиболее значимых качеств связаны генетически с вредными и деструктивными. Сотрудничество с членами группы может сочетаться с агрессивностью по отношению к чужакам, творческий дар – с жаждой владеть и властвовать, атлетическое усердие – со склонностью к насильственным действиям, и так далее. В предельных случаях такие сочетания – следствие плейотропизма, когда один набор генов управляет более чем одной фенотипической характеристикой. Если планируемое общество, создание которого кажется неизбежным в 21 веке, должно будет сознательно проводить своих членов мимо от тех стрессов и конфликтов, которые подставили деструктивные фенотипы под “нож Дарвина”, с ними могут вырождаться другие фенотипы. В этом, чисто генетическом, смысле, социальный контроль лишит человека гуманности.
Похоже, что наша автокаталитическая социальная эволюция зажала нас в специфическом канале, который ранние гоминиды, что все ещё внутри нас, могут не приветствовать. Чтобы поддерживать вид неограниченно, мы должны двигаться в направлении тотального знания, буквально до уровня отдельного нейрона и гена. Когда мы достигнем решительного прогресса в понимании самих себя в механистических терминах, и социальные науки достигнут полного расцвета, с результатом всего этого может оказаться трудно смириться. Поэтому, похоже, уместно закончить эту книгу тем же, с чего она начата – с недоброго пророчества Альбера Камю:
Мир, который можно объяснить даже с плохими доводами – это наш привычный мир. С другой стороны, в этой вселенной, лишенной иллюзий и светил, человек чувствует себя лишним. Его изгнание – без средств к излечению, ибо он лишен памяти об утерянном доме и надежд на землю обетованную.
Это, к сожалению, правда. Но у нас всё ещё есть другая сотня лет.
Библиография
- Adams, R. McC. 1966. The evolution of urban society: early Mesopotamia and prehispanic Mexico. Aldine Publishing Co., Chicago, xii + 191 pp.
- Alexander, R. D. 1971. The search for an evolutionary philosophy of man. Proceedings of the Royal Society of Victoria, 84(1): 99-120.
- Bartholomew, G. A., and J. B. Birdsell. 1953. Ecology and the protohominids. American Anthropologist, 55: 481-498.
- Bigelow, R. 1969. The dawn warriors: man’s evolution toward peace. Atlantic Monthly Press, Little, Brown, Boston. xi + 277 pp.
- Birdwhistle, R. L. 1970. Kinesics and context: essays on body motion and communication. University of Pennsylvania Press, Philadelphia, xiv + 338 pp.
- Boserup, Ester. 1965. The conditions of agricultural growth. Aldine Publishing Co., Chicago. 124 pp.
- Brannigan, C. R., and D. A. Humphries. 1972. Human non-verbal behaviour, a means of communication. In N. Blurton Jones, ed. (q.v.). Ethological studies of child behaviour, pp. 37–64.
- Brown, R. 1973. A first language: the early stages. Harvard University Press, Cambridge, xxii + 437 pp.
- Campbell, D. T. 1972. On the genetics of altruism and the counterhedonic components in human culture. Journal of Social Issues, 28(3): 21-37.
- Carneiro, R. L. 1970. A theory of the origin of the state. Science, 169: 733-738.
- Chagnon, N. A. 1968. Yanomamo: the fierce people. Holt, Rinehart and Winston, New York. xviii + 142 pp.
- Chance, M. R. A. 1962. Social behaviour and primate evolution. In M. F. Ashley Montagu, ed.. Culture and the evolution of man, pp. 84-130. Oxford University Press, New York. xiii + 376 pp.
- Chomsky, N. 1957. Syntactic structures. Mouton, The Hague. 118 pp.
- Chomsky, N. 1972. Language and mind, enlarged ed. Harcourt, Brace, Jovanovich, New York. xii + 194 pp.
- Count, E. W. 1958. The biological basis of human sociality. American Anthropologist, 60(6): 1049-1085.
- Crystal, D. 1969. Prosodic systems and intonation in English. Cambridge University Press, London, viii + 381 pp.
- Dahlberg, G. 1947. Mathematical methods for population genetics. S. Karger, New York. 182 pp.
- Darlington, C. D. 1969. The evolution of man and society. Simon and Schuster, New York. 753 pp.
- Dart, R. A. 1949. The predatory implemental technique of Australopithecus. American Journal of Physical Anthropology, n.s. 7: 1-38.
- Dart, R. A. 1956. Cultural status of the South African man-apes. Report of the Smithsonian Institution, Washington, D. C., 1955, pp. 317-338.
- Darwin, C. 1871. The descent of man, and selection in relation to sex, 1 vols. Appleton, New York. Vol. 1: vi + 409 pp.; vol. 2: viii + 436 pp.
- Denes, P. B., and E. N. Pinson. 1973. The speech chain: the physics and biology of spoken language, rev. ed. Anchor Press, Doubleday, Garden City, N. Y. xviii + 217 pp.
- Dobzhansky, T. 1963. Anthropology and the natural sciences – the problem of human evolution. Current Anthropology, 4: 138, 146-148.
- Eshel, I. 1972. On the neighbor effect and the evolution of altruistic traits. Theoretical Population Biology, 3(3): 258-277.
- Etkin, W. 1954. Social behavior and the evolution of man's mental faculties. American Naturalist, 88(840): 129-142.
- Flannery, K. V. 1972. The cultural evolution of civilizations. Annual Review of Ecology and Systematics, 3: 399-426.
- Fodor, J., and M. Garrett. 1966. Some reflections on competence and performance. In J. Lyons and R. J. Wales, eds., Psycholinguistic papers, pp. 133–163. Edinburgh University Press, Edinburgh. 243 pp.
- Fox, M. W. 1972. Socio-ecological implications of individual differences in wolf litters: a developmental and evolutionary perspective. Behaviour, 46(3,4): 298-313.
- Fox, R. 1971. The cultural animal. In J. F. Eisenberg and W. S. Dillon, eds., Man and beast: comparative social behavior, pp. 263-296. Smithsonian Institution Press, Washington, D. C. 401 pp.
- Fox, R. 1972. Alliance and constraint: sexual selection in the evolution of human kinship systems. In B. G. Campbell, ed., Sexual selection and the descent of man 1871-1971, pp. 282-331. Aldine Publishing Co., Chicago. x + 378 pp.
- Freedman, D. G. 1974. Human infancy: an evolutionary perspective. Lawrence Eribaum, Hillsdale, N. J. xii + 212 pp.
- Freedman, D. G. 1979. Human sociobiology. Free Press, New York. 188 pp.
- Friedrichs, R. W. 1970. A sociology of sociology. Free Press, Collier-Macmillan, New York. xxxiv + 429 pp.
- Goffman, E. 1959. The presentation of self in everyday life. Doubleday Anchor Books, Doubleday, Garden City, N. Y. xvi + 259 pp.
- Goffman, E. 1961. Encounters: two studies in the sociology of interaction. Bobbs-Merrill, Indianapolis. 152 pp.
- Grant, E. C. 1969. Human facial expression. Man, 4(4): 525-536.
- Haldane, J. B. S. 1932. The causes of evolution. Longmans, Green, London, vii + 234 pp. (Reprinted as a paperback, Comell University Press, Ithaca, N. Y., 1966. vi + 235 pp.)
- Hardin, G. 1972. Population skeletons in the environmental closet. Bulletin of the Atomic Scientists, 28(6) (June): 37-41.
- Helm, June. 1968. The nature of Dogrib socioterritorial groups. In R. B. Lee and I. DeVore, eds. (q.v.), Man the hunter, pp. 118–125.
- Herrnstein, R. J. 1971b. I. Q. Atlantic Monthly, 228(3) (September): 43–64.
- Hill, Jane H. 1972. On the evolutionary foundations of language. American Anthropologist, 74(3): 308-317.
- Homans, G. C. 1961. Social behavior: its elementary forms. Harcourt, Brace & World, New York. xii + 404 pp.
- Hooff, J. A. R. A. M. van. 1972. A comparative approach to the phylogeny of laughter and smiling. In R. A. Hinde, ed., Non-verbal communication, pp. 209-241. Cambridge University Press, Cambridge, xiii + 423 pp.
- Howells, W. W. 1973. Evolution of the genus Homo. Addison-Wesley, Reading, Mass. 188 pp.
- Hutchinson, G. E. 1959. A speculative consideration of certain possible forms of sexual selection in man. American Naturalist, 93(869): 81-91.
- Inkeles, A. 1964. What is sociology? An introduction to the discipline and profession. Prentice-Hall, Englewood Cliffs, N. J. viii + 120 pp.
- Jolly, C. J. 1970. The seed-eaters: a new model of hominid differentiation based on a baboon analogy. Man, 5(1): 5-26.
- Jones, N. Blurton ed. 1969. Ethological studies of child behaviour.
- Keith, A. 1949. A new theory of human evolution. Philosophical Library, New York. x + 451 pp.
- Kohlberg, L. 1969. Stage and sequence: the cognitive-developmental approach to socialization. In D. A. Goslin, ed., Handbook of socialization theory and research, pp. 347-480. Rand McNally Co., Chicago, xiii + 1182 pp.
- Kortlandt, A. 1972. New perspectives on ape and human evolution. Stichting voor Psychobiologie, Universiteit van Amsterdam, The Netherlands. 100 pp.
- Kummer, H. 1971. Primate societies: group techniques of ecological adaptation. Aldine-Atherton, Chicago. 160 pp.
- Kurtén, B. 1972. Not from the apes. Vintage Books, Random House, New York. viii + 183 pp.
- Lee, R. B., and I. DeVore, eds. 1968. Man the hunter. Aldine Publishing Co., Chicago, xvi + 415 pp.
- Leibowitz, Lila. 1968. Founding families. Journal of Theoretical Biology, 21(2): 153–169.
- Lemer, I. M. 1968. Heredity, evolution, and society. W. H. Freeman, San Francisco, xviii + 307 pp.
- Lenneberg, E. H. 1967. Biological foundations of language. John Wiley & Sons, New York. xviii + 489 pp.
- Lenneberg, E. H. 1971. Of language knowledge, apes, and brains. Journal of Psycholinguistic Research, 1(1): 1-29.
- Lenski, G., and Jean Lenski. 1970. Human societies: a macro-level introduction to sociology. McGraw-Hill Book Co., New York. xvi + 515 pp.
- LeVine, R. A., and D. T. Campbell. 1972. Ethnocentrism: theories of conflict, ethnic attitudes, and group behavior. John Wiley & Sons, New York. x + 310 pp.
- Lévi-Strauss, C. 1949. Les structures elementaires de la parenle. Presses Universitaires de France, Paris. xiv + 639 pp. (The elementary structures of kinship, rev. ed., trans. by J. H. Bell and J. R. von Stunner and ed. by R. Needham, Beacon Press, Boston, 1969. xlii + 541 pp.
- Lieberman, P. 1968. Primate vocalizations and human linguistic ability. Journal of the Acoustic Society of America, 44: 1574-1584.
- Lieberman, P., E. S. Crelin, and D. H. Klatt. 1972. Phonetic ability and related anatomy of the newbom and adult human. Neanderthal man, and the chimpanzee. American Anthropologist, 74(3): 287-307.
- Lyons, J. 1972. Human language. InR. A. Hinde, ed. (q.v.), Non-verbal communication, pp. 49–85.
- Marler, P. R. 1965. Communication in monkeys and apes. In I. DeVore, ed.. Primate behavior: field studies of monkeys and apes, pp. 544-584. Holt, Rinehart and Winston, New York. xiv + 654 pp.
- Martin, N. G., L. J. Eaves, and H. J. Eysenck. 1977. Genetical, environmental and personality factors in influencing the age of first sexual intercourse in twins. Journal of Biosocial Science, 9(1): 91-97.
- Martin, P. S. 1966. Africa and Pleistocene overkill. Nature, London, 212(5060): 339-342.
- Maslow, A. H. 1954. Motivation and personality. Harper, New York. 411 pp.
- Maslow, A. H. 1972. The farther reaches of human nature. Viking Press, New York. xxii + 423 pp. Masters, R. D. 1970. Genes, language, and evolution. Semiotica, 2(4): 295-320.
- Masters, R. D. 1970. Genes, language, and evolution. Semiotica, 2(4): 295-320.
- Masters, W. H., and Virginia E. Johnson. 1966. Human sexual response. Little, Brown, Boston, xiii + 366 pp.
- Mattingly, I. G. 1972. Speech cues and sign stimuli. American Scientist, 60(3): 327-337.
- McEvedy, C. 1967. The Penguin atlas of ancient history. Penguin Books, Baltimore Md. 96 pp.
- Miller, G. A., E. Galanter, and K. H. Pribram. 1960. Plans and the structure of behavior. Henry Holt, New York. xii + 226 pp.
- Morris, D. 1962. The biology of art. Alfred Knopf, New York. 176 pp.
- Morris, D. 1967. The naked ape: a zoologist's study of the human animal. McGraw-Hill Book Co., New York. 252 pp.
- Morton, N. E. 1969. Human population structure. Annual Review of Genetics, 3: 53–74.
- Napier, J. R., and P. H. Napier. 1967. A handbook of living primates. Academic Press, New York. xiv + 456 pp.
- Ogburn, W. F. and M. Nimkoff. 1958. Sociology, 3d ed. Houghton Mifflin Co., Boston, x + 756 pp.
- Parsons, P. A. 1967. The genetic analysis of behaviour. Methuen, London, x + 174 pp.
- Patterson, O.1967. The sociology of slavery: an analysis of the origins, development and structure of Negro slave society in Jamaica. Fairleigh Dickinson University Press, Cranbury, N. J. 310 pp.
- Pfeiffer, J. E. 1969. The emergence of man. Harper & Row, New York. xxiv + 477 pp.
- Pilbeam, D. 1972. The ascent of man: an introduction to human evolution. Macmillan Co., New York. x + 207 pp.
- Rabb, G. B., J. H. Woolpy, and B. E. Ginsburg. 1967. Social relationships in a group of captive wolves. American Zoologist, 7(2): 305-311.
- Rappaport, R. A. 1971. The sacred in human evolution. Annual Review of Ecology and Systematics, 2: 23-44.
- Rawls, J. 1971. A theory of justice. Belknap Press of Harvard University Press, Cambridge, xvi + 607 pp.
- Rensky, M. 1966. The systematics of paralanguage. Travaux linguistiques de Prague, 2: 97–102.
- Reynolds, V. 1968. Kinship and the family in monkeys, apes and man. Man, 3(2): 209–233.
- Rousseau, M. 1971. Un machairodonte dans 1’art Aurignacien? Mammalia, 35(4): 648-657.
- Schaller, G. B., and G. R. Lowther. 1969. The relevance of carnivore behavior to the study of early hominids. Southwestern Journal of Anthropology, 25(4): 307-341.
- Scott, J. F. 1971. Intemalization of norms: a sociological theory of moral commitment. Prentice-Hall, Englewood Cliffs, N. J. xviii + 237 pp.
- Simons, E. L., and P. C. Ettel. 1970. Gigantopithecus. Scientific American, 222(1) (January): 76-85.
- Sipes, R. G. 1973. War, sports and aggression: an empirical test of two rival theories. American Anthropologist, 75(1): 64-86.
- Slobin, D. 1971. Psycholinguistics. Scott, Foresman, Glen-view, 111. xii + 148 pp.
- Stent, G. S., ed. 1978. Morality as a biological phenomenon. Dahlem Workshop. Abakon Verlagsgesellschaft, Berlin. 323 pp.
- Thorpe, W. H. 1972. The comparison of vocal communication in animals and man. In R. A. Hinde, ed. (q.v.), Non-verbal communication, pp. 27–47. Cambridge University Press, Cambridge, xiii + 423 pp.
- Tiger, L., and R. Fox. 1971. The imperial animal. Holt, Rinehart and Winston, New York. xi + 308 pp.
- Tobias, P. V. 1973. Implications of the new age estimates of the early South African hominids. Nature, London, 246(5428): 79-83.
- Trivers, R. L. 1971. The evolution of reciprocal altruism. Quarterly Review of Biology, 46(4): 35-57.
- Washburn, S. L. 1970. Comment on: “A possible evolutionary basis for aesthetic appreciation in men and apes”. Evolution, 24(4): 824-825.
- Washburn, S. L., and F. C. Howell. 1960. Human evolution and culture. In S. Tax, ed., Evolution after Darwin, vol. 2, Evolution of man, pp. 33-56. University of Chicago Press, Chicago, viii + 473 pp.
- Washburn, S. L., and Virginia Avis. 1958. Evolution of human behavior. In Anne Roe and G. G. Simpson, eds., Behaviour and evolution, pp. 421-436. Yale University Press, New Haven, Conn. vii + 557 pp.
- Washburn, S. L., ed. 1961. Social life of early man. Viking Fund Publications in Anthropology no. 31. Aldine Publishing Co., Chicago, ix + 299 pp.
- Weinrich, J. D. 1976. Human reproductive strategy: the importance of income unpredictability, and the evolution of non-reproduction. Ph.D. thesis. Harvard University, Cambridge. 231 pp.
- White, H. C. 1970. Chains of opportunity: system models of mobility in organizations. Harvard University Press, Cambridge, xvi + 418 pp.
- Whiting, J. W. M. 1968. Discussion, “Are the hunter-gatherers a cultural type?” In R. B. Lee and I. DeVore, eds. (q.v.), Man the hunter, pp. 336-339.
- Wilmsen, E. N. 1973. Interaction, spacing behavior, and the organization of hunting bands. Journal of Anthropological Research, 29(1): 1-31.
- Wilson, E. O. 1971a. The insect societies. Belknap Press of Harvard University Press, Cambridge, x + 548 pp.
- Wilson, E. O. 1972a. On the queerness of social evolution. Bulletin of the Entomological Society of America, 19(1): 20-22.
- Wilson, E. O. 1978. On human nature. Harvard University Press, Cambridge, xii + 260 pp.
- Wilson, R. S. 1978. Synchronies in mental development: an epigenetic perspective. Science, 202: 939-948.