
- Орангутаны научились использовать орудия с выгодой для себя [2019-03-22]
- Мозг собак отличил настоящие слова от тарабарщины [2019-03-22]
- Какаду Гоффина выклевали палочки из картонки и использовали их в быту [2019-03-22]
- Орангутанов уличили в создании крючков [2019-03-22]
- Любовь самок гуппи к ярким самцам объяснили генами и освещением [2019-03-22]
- Орангутаны рассказывают друг другу о прошлом [2019-03-22]
- Блеск и нищета этологии [2018-05-03]
- Почему хозяева похожи на своих собак? [2018-03-15]
- Интервью с Анатолием Протопоповым [2019-02-13]
- Цирки: развлечение vs мучение [2018-12-15]

Н. Тинберген
Перевод с английского
Ю.Л. Амченкова
Под редакцией
Акад. РАН П.В. Симонова
Social Behaviour In Animals
With Special Reference To Vertebrates By N.Tinbergen
Lectures In Animal Behaviour In The University Of Oxford
First published in 1953
ПРЕДИСЛОВИЕ РЕДАКТОРА ПЕРЕВОДА
Предлагаемую вниманию читателей книгу Николаса Тинбергена (1907 – 1988) “Социальное поведение животных” можно с полным основанием считать одним из классических руководств, посвященных относительно новой области современных биологических знаний, – этологии. Именно в этом качестве книга, многократно переиздававшаяся с 1953 г., не утратила своей познавательной ценности для русскоязычной аудитории.
Признанием значения этологии как специальной отрасли естествознания явилось присуждение автору книги совместно с Карлом фон Фришем и Конрадом Лоренцем Нобелевской премии по физиологии и медицине за 1973 г. Этология – наука о сложных формах поведения животных в естественной для них среде обитания. Такого рода исследования в значительной мере построены на наблюдениях, но отнюдь не сводятся к ним, обладая всеми характерными признаками научного подхода, т.е. формулировкой гипотез, подлежащих тщательной проверке.
Н. Тинберген подробно перечисляет методы, используемые этологами для получения достоверных знаний о закономерностях и механизмах поведения. Во-первых, это – повторные наблюдения, уточняющие реальность существования и детали ранее зарегистрированных фактов. Они ведутся с помощью разнообразных укрытий, средств дистанционного слежения, фото- и киносъемки. Полученные таким образом данные проверяются в экспериментах, где, например, естественные цветы заменяются различно окрашенными чашками с сиропом, а живые существа – макетами С окраской, характерной для видоспецифичных стимулов – “релизеров”, способных вызвать генетически детерминированную реакцию. В необходимых случаях эксперимент организуется в условиях относительной полусвободы животных: в зоопарках, аквариумах и океанариях. Таким образом, современный этологический эксперимент весьма существенно отличается от любознательности непрофессиональных любителей природы и позволяет говорить об этологии как о науке в общепринятом смысле слова.
Социальное поведение Н. Тинберген определяет как взаимодействие между особями одного и того же вида, специально подчеркнув, что не всякая групповая активность будет социальной. Совместный полет бабочек к источнику света или общее бегство животных от лесного пожара не могут быть названы “социальным поведением”. Биологическая ценность последнего в том, что оно позволяет решать адаптивные задачи, непосильные для отдельно взятой особи. Только точная и взаимная синхронизация действий брачных партнеров приводит к оплодотворению. Трудно себе представить выживание беспомощного молодняка без родительской заботы о нем. Зоосоциальные сигналы опасности и совместное нападение на врага обеспечивают эффективную защиту от охотящихся хищников, а внутригрупповая иерархия исключает негативные последствия схваток при каждом дележе пищи.
Длительный процесс эволюции сделал проявления социального поведения внешне столь целесообразными, что они кажутся разумными поступками и позволяют предполагать у животных некое подобие рассудочной деятельности. Примером может служить замена брачных, территориальных и иерархических драк демонстрацией угрожающих действий или поз подчинения. Однако тщательный анализ обнаруживает их врожденную запрограммированность. Так, крик, служащий коммуникативным сигналом опасности, птица издает и в полном одиночестве, когда ей некого предупреждать о возникшей угрозе.
Поскольку свои исследования Н. Тинберген проводил на птицах, рыбах и насекомых, он имел дело в основном с инстинктивными, врожденными формами социального поведения. Но и на этом уровне автор не мог не столкнуться с примерами этологической пластичности, а также взаимодействия врожденных и приобретенных свойств.
Дело в том, что реализация генетически запрограммированных реакций подчас решающим образом зависит от текущего функционального состояния животного. Например, реакция на яйцо (поведение насиживания) определяется гормональным статусом птицы, содержанием в ее крови секретируемого гипофизом гормона – пролактина. Важен и возраст животного. Выдающемуся советскому физиологу Л. А. Орбели принадлежит стройная, всесторонне аргументированная концепция постнатального дозревания врожденных безусловных рефлексов под влиянием и при взаимодействии с условными.
Многочисленные примеры вмешательства условных рефлексов в реализацию безусловных приводит в своей книге и Н. Тинберген. Когда паре цихловых рыб подменили мальков, рыбы стали заботиться о “приемышах”, относящихся к другому виду, но одновременно питаться мальками своего. При следующем нересте они съели собственных мальков. Многие животные (особенно млекопитающие) реагируют на видоспецифичные “релизеры” только знакомой особи, а пчелы и шмели начинают собирать нектар только с определенного вида растений. Еще более сложные функциональные перестройки наблюдаются в сообществах с узкой специализацией членов. Если из улья удалить пчел-сборщиц нектара, то за ним станут летать те особи, которые раньше были заняты кормлением личинок. Заметим, что в исследование взаимодействия врожденных и индивидуально приобретаемых факторов поведения большой вклад внесли советские ученые: физиолог П.К. Анохин, генетик Д.К. Беляев, зоолог М.С. Гиляров и др.
Н. Тинберген завершает свое увлекательное изложение кратким очерком эволюции зоосоциального поведения. Он справедливо полагает, что поражающие нас своей кажущейся целесообразностью поведенческие акты вначале носили случайный характер, но позднее были закреплены естественным отбором. Например, материалом для формирования движений - “релизеров” могли послужить проявления смещенной активности, возникающей при конфликте мотиваций. Так, при одновременной активизации половой потребности и агрессивности птица начинает яростно щипать траву, т. е. осуществлять действие, характерное для пищедобывательного поведения, хотя пищевая мотивация в данном случае отсутствует.
Что касается эволюционного происхождения альтруистического поведения, то его основу составляет так называемый отбор родичей, при котором гибель отдельных особей обеспечивает сохранение генов близкородственных им организмов. Вот почему Говорить об альтруизме в человеческом смысле допустимо только в том случае, когда речь идет о помощи “неродным” существам. Согласно современным представлениям, альтруистическое доведение у людей определяется двумя основными мотивациями:
Механизмом сопереживания, сочувствия и потребностью следовать этическим нормам, принятым в обществе. На примере альтруизма мы хотим подчеркнуть ту величайшую осторожность, которую следует проявлять при сопоставлении социального поведения животных и человека, наделенного сознанием и феноменом культурного (негенетического) наследования. Об этих принципиальных различиях неоднократно упоминает в своей книге и Н. Тинберген. Сказанное ни в коей мере не умаляет значения этологических концепций не только для науки о поведении животных, но и для человековедения, для проникновения в биологические корни антропосоциогенеза. Вот почему нам хочется закончить свое предисловие словами И. П. Павлова:
“Нет никакого сомнения, что систематическое изучение фонда прирожденных реакций животного чрезвычайно будет способствовать пониманию нас самих и развитию в нас способности к личному самоуправлению” (Павлов И. П. Двадцатилетний опыт изучения высшей нервной деятельности (поведения) животных. М.: Наука, 1973, с. 240).
П. В. Симонов
Эта книга не ставит перед собой целью исчерпывающим образом перечислить известные факты. Ее задача состоит скорее в биологическом подходе к феномену социального поведения. Такой подход четко прослеживается в исследованиях Лоренца. Для него характерны упор на неоднократные и тщательные наблюдения за колоссальным разнообразием происходящих в природе социальных взаимодействий, на одинаково пристальное внимание к трем важнейшим биологическим проблемам: функции, причинности, эволюции, — упор на правильную последовательность описания явления и его качественного и количественного анализа и, наконец. на непрерывное переосмысление данных.
Особенности этого подхода наряду с недостатком места определили содержание книги. Ограниченный ее объем привел к отказу от большого числа описаний. Например, здесь не обсуждается объемистая работа Дегенера о различных типах агрегации животных. Не рассматриваются в деталях и высокоспециализированные “государства” общественных насекомых, поскольку существует ряд прекрасных обзоров, посвященных исключительно этому вопросу.
Природа выбранного подхода привела к принципиальному отличию книги от других работ по социальному поведению. С одной стороны, я кратко коснулся некоторых проблем, которые гораздо подробнее рассмотрены моими коллегами. Так, публикации Алли посвящены в основном различным преимуществам, получаемым животными от жизни в стае. в них мало говорится о феноменах, на которых основано социальное взаимодействие, а если они и упоминаются, внимание концентрируется исключительно на “порядке клевания” — интересном, но не главном аспекте социальной организации. Другие специалисты, на мой взгляд, незаслуженно большую роль приписывают передаче пищи от одной особи другой; хотя и признано, что это входит в число факторов развития определенных социальных отношений, речь идет опять же всего лишь об одном из элементов широкого комплекса феноменов. Наконец, накоплено огромное количество часто не связанных между собой аналитических данных, полученных в лабораторных условиях, о которых на сегодняшний день невозможно сказать, как они соотносятся с нормальной жизнью исследованных видов.
С другой стороны, я придавал огромное значение формулировке важнейших проблем, показу их связей между собой и с другими, более специальными и подчиненными проблемами. Эта задача вместе с необходимостью описания многих новых фактов, выяснившихся в ходе “натуралистических” наблюдений, и также первых попыток их качественного анализа потребовала много места, Кроме того, мне хотелось сформулировать и обсудить некоторые новые теории, которые мне кажутся важными в связи с их большой эвристической ценностью. Так, достаточно детально представлены значение внутривидовых драк, причинно-следственные связи во время угрожающего поведения и ухаживания, функции релизеров и другие проблемы, пониманию которых во многом способствовал упомянутый новый подход; я попытался найти их истинное место в сложной системе устоявшихся представлений о предмете.
Я старался излагать свои мысли так, чтобы за ними могли без труда и с интересом следить даже непрофессионалы. Надеюсь, это пойдет на пользу будущим исследованиям, поскольку, на мой взгляд, наша молодая наука может получить много полезного от работ любителей.
Я глубоко признателен доктору Майклу Аберкромби и Дезмонду Моррису за их ценные критические замечания и просмотр текста, доктору Л. Тинберген за ряд сделанных иллюстраций, а также издательству “Oxford University Press” за разрешение использовать здесь несколько рисунков из моей книги “The Study of Instinct”. Большое спасибо также доктору Хью Котту за разрешение воспроизвести рис. 61 и доктору Брайану Робертсу за возможность использовать его прекрасный снимок пингвинов, приведенный на вкладке 5.
ГЛАВА 1
СУТЬ ВОПРОСА
Если скворцов, живущих стаями, мы уверенно называем социальными животными, а сокол-сапсан, охотящийся зимой над заливом, птица как будто одиночная, значит, термин “социальный” указывает на то, что речь идет явно не об одной особи. Их не обязательно должно быть много. “Социальными” я буду в дальнейшем называть многие особенности поведения, проявляемые всего парой взаимодействующих индивидов.
Однако социальность свойственна не всем агрегациям животных. Когда летней ночью тысячи насекомых собираются вокруг лампы, их поведение не обязательно социальное. Возможно, они прилетели поодиночке, и их скопление в данном месте—явная случайность, результат того, что каждая особь привлекается одним и тем же—светом. Однако скворцы, совершающие зимними вечерами свои поразительные воздушные маневры, перед тем как спуститься на ночевку,—действительно реагируют друг на друга. Они следуют один за другим в таком правильном порядке, что может даже возникнуть мысль о сверхчеловеческой способности к общению. Значит, совместные действия, основанные на взаимном реагировании,—еще одна особенность социального поведения. Этим зоосоциология отличается от фитосоциелогии, изучающей все феномены, возникающие при совместном произрастании растений, независимо от того, влияют они друг на друга или просто оказались вместе в результате одинаковой реакции на одни и те же внешние факторы.
Влияние, оказываемое социальными животными друг на друга, не сводится к простому взаимному привлечению. Агрегация обычно лишь прелюдия к более тесному сотрудничеству, т. е. совместному совершению тех или иных действий. В случае скворцов такое сотрудничество очевидно—они вместе летают, выполняя одинаковые повороты, некоторые могут издавать предупреждающие крики, на которые другие реагируют, бывает, что они объединяются, чтобы отразить атаку ястреба-перепелятника или сокола-сапсана, когда сбившись плотной массой, поднимаются над хищником. За совместными полетами самца и самки в сезон размножения может следовать длительный период тесного и сложного сотрудничества во время спаривания, строительства гнезда, высиживания и выращивания птенцов.
Таким образом, изучение социального поведения—это изучение сотрудничества между особями. Их может быть две или более. В стае скворцов это сотрудничество иногда объединяет тысячи особей.
Говоря о сотрудничестве, мы всегда явно или неявно подразумеваем его цель, т. е. предполагаем, что оно для чего-то служит. Эта проблема “биологического смысла”, или “функции” жизненных процессов,— одна из наиболее популярных среди исследователей. Она возникает при изучении физиологии особи и каждого из ее органов. С другой стороны, на более высоком уровне интеграции она существует и в зоосоциологии. Если физик или химик не стремится понять цели исследуемых феноменов, биолог должен это делать. Естественно, “цель” понимается здесь в узком смысле слова. Это не значит, что биолог больше озабочен вопросом, зачем вообще нужна жизнь, чем физик — конечной целью существования материи и движения. Однако сама природа живого, его неустойчивость заставляют спрашивать: почему организмы не разрушаются под действием присутствующих повсюду враждебных влияний среды? Как им удается выживать, поддерживать свое существование и производить себе подобных? Цель, задача или смысл биологических процессов в этом узком понимании— сохранение особи, группы и вида. Сообщество индивидов должно поддерживать свое существование, защищаться от распада не меньше, чем отдельный организм, который, как следует из самого этого термина, является сообществом собственных частей— органов, а также частей органов, частей частей органов и т. д. Если физиолог задается вопросом, как индивиду, органу или клетке удается сохранить себя в ходе организованного взаимодействия своих составляющих, зоосоциолог должен спрашивать себя: как членам группы—индивидам—удается сохранять группу.
В этой главе я прежде всего для ясности приведу ряд примеров групповой жизни нескольких видов животных. Затем в следующих главах проанализирую, какие функдии особей при социальном поведении приносят пользу другим индивидам или группе в целом. После этого я рассмотрю, каким образом организовано сотрудничество. Два этих аспекта, т.е. 'функция и причинно-следственная обусловленность социального поведения, будут обсуждаться применительно к нескольким его типам: взаимодействию половых партнеров, семейной и групповой жизни, дракам между особями. В результате шаг за шагом мы подойдем к социальным структурам. Поскольку такие структуры почти всегда временные, придется рассмотреть, как они возникают. Наконец, следует попытаться понять пути развития современных типов социальной организации в ходе долгой эволюции организмов.
СЕРЕБРИСТАЯ ЧАЙКА (LARUS ARGENTATUS)
Всю осень и зиму серебристые чайки живут стаями. Стайно питаются, мигрируют, спят. Если наблюдать изо дня в день за серебристыми чайками, занимающимися поиском пищи, можно заметить, что, как правило, их собирает вместе не общая реакция на внешний фактор типа обильной пищи. Одна из известных мне групп чаек обычно кормилась земляными червями на лугах, причем день—на одном лугу, другой—на другом. В обоих этих местах червей было предостаточно, и ничто не говорило о том, что чайки меняют место кормежки из-за недостатка пищи. В самом деле, существенно сократить популяцию земляных червей очень нелегко! Когда отдельные чайки прибывали на кормовой участок, они всегда собирались вместе и не садились поодиночке на удаленные от стаи места луга. Их привлекали именно другие чайки.
Птицы в стае реагировали друг на друга различным образом. Если подойти к ним слишком близко, некоторые чайки перестают питаться, вытягивают шею и пристально смотрят на вас. Вскоре то же самое делают другие. Наконец, вся стая стоит, уставившись на пришельца. Затем одна из чаек может издать крик тревоги — ритмичное “га-га-га”—и тут же взлететь. За ней немедленно последуют другие, и в результате вся стая снимется с места. Реакция почти одновременная. Конечно, не исключено, что это результат их одновременной реакции на вас как на внешний фактор, запускающий такое поведение. Однако довольно часто, например когда вы подкрадываетесь к ним под покрывалом, обнаружить вас способны только одна-две птицы, после чего видно, как их поведение—вытягивание шеи, крик или внезапный взлет—влияет на других, которые, возможно, и не заметили опасности сами.
Весной вся стая прилетает на гнездовые участки в песчаных дюнах. Когда птицы, покружив некоторое время в воздухе, опускаются на землю, они разделяются на пары, занимающие отдельные территории в пределах колонии. Однако пары образуют не все особи, многие объединяются, так сказать, в “клубы”. Продолжительное изучение меченых особей показало, что новые пары формируются в таких клубах, причем инициативу здесь берут на себя самки. Оставшаяся без партнера самка приближается к самцу особым образом. Она втягивает шею, направляет клюв вперед и слегка вверх, а затем, расположив тело горизонтально, медленно кружит вокруг выбранного самца. Тот может реагировать двумя способами: либо начинает с важным видом оборачиваться и нападать на других самцов, либо издает протяжный крик и отходит вместе с самкой. Тогда она часто начинает выпрашивать у него корм, своеобразно подергивая головой. Самец реагирует на такое выпрашивающее поведение, отрыгивая часть проглоченной пищи, которую самка жадно съедает (рис. 1). В начале сезона размножения это может быть просто “флирт”, не завершающийся серьезной связью. Однако обычно птицы в таких парах привязываются друг к другу, что ведет к заключению прочного союза. Когда он возник, делается следующий шаг: поиск места для гнезда. Птицы покидают клуб и выбирают собственную территорию в пределах занятого колонией пространства. Здесь они начинают строить гнездо. Оба партнера собирают гнездовой материал и переносят его к облюбованному месту, где, усаживаясь по очереди на землю, выкапывают ногами что-то вроде мелкого колодца, который выстилают травой и мхом.
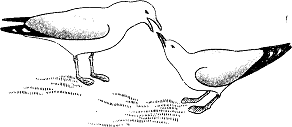
Рис. 1. Самец серебристой чайки (слева) готовится кормить самку
Раз или два в день птицы спариваются. Этому всегда предшествует долгая церемония. Оба партнера начинают подергивать головой, как будто выпрашивают корм. Разница с “кормлением ухаживания” заключается в том, что такие движения делают и самец, и самка. Спустя некоторое время самец начинает постепенно вытягивать шею, вскоре после чего вспрыгивает на самку. Спаривание заключается в неоднократном соприкосновении клоак партнеров.
Одновременно с образованием пар, строительством гнезда, ухаживанием и спариванием можно наблюдать и другие типы поведения, в частности драки самцов. Уже внутри клуба агрессивность самца может быть настолько высокой, что он разгоняет всех находящихся рядом чаек. Обосновавшись на своей гнездовой территории, он становится совершенно нетерпимым к нарушителям ее границы, нападая на каждого подошедшего слишком близко самца. Обычно настоящей атаки не происходит: одной лишь угрозы часто достаточно для того, чтобы пришелец удалился. Существуют три типа угроз. Самая мягкая форма—это “вертикальная угрожающая поза”: самец вытягивает шею, направляет клюв вниз и иногда приподнимает крылья (рис. 2). Приняв такое положение, он очень скованной походкой направляется к чужаку, все его мышцы напряжены. Более сильное выражение враждебных намерений — “дерганье травы”. Самец подходит довольно близко к противнику, нагибается, сердито клюет землю, хватает клювом пучок травы, мха или корней и вырывает его. Когда самец и самка сталкиваются с соседней парой вместе, они демонстрируют третий тип угрозы—“задыхающийся”, т.е. приседают, опускают грудь и направляют вниз клюв с опущенной подъязычной костью, что придает им весьма любопытное “выражение лица”. Затем они делают ряд незавершенных как бы клюющих движений, направленных в сторону земли, сопровождая их ритмичным хриплым воркованием.
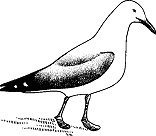
Рис. 2. Вертикальная поза угрозы самца серебристой чайки
Все эти действия явно производят впечатление на других чаек, которые понимают их агрессивный характер и часто отступают.
Когда яйца отложены, партнеры по очереди их насиживают.
Здесь опять же хорошо заметно сотрудничество между чайками. Партнеры никогда не оставляют яйца без присмотра; если один сидит на них, второй может искать пищу в нескольких милях от гнезда. Когда он возвращается, насиживающая птица ожидает, пока партнер дойдет до гнезда. Свое приближение он сопровождает особыми движениями и криками: обычно издает продолжительный “мяукающий” призыв, часто приносит с собой немного гнездового материала. Затем сидящая птица поднимается, а вторая занимает ее место.
Заботу о яйцах можно было бы отнести к социальному поведению, поскольку с момента откладки они считаются особями. Обычно мы не рассматриваем такие односторонние отношения как истинно социальные, но не следует забывать, что яйцо, хотя и не движется, обеспечивает особые стимулы, глубоко влияющие на птицу-родителя.
Однако как только выводятся птенцы, отношения между родителями и потомством становятся, бесспорно, взаимными. Сначала птенцы в основном пассивно обогреваются, но спустя несколько часов начинают выпрашивать корм. Когда родитель дает им возможность подняться на ноги, они делают ряд клюющих движений, направленных к его клюву. Чайка отрыгивает пищу, т. е. полупереваренную рыбу или краба, или комок земляных червей, зажимает кусочек этой массы концом клюва и терпеливо предлагает его птенцам (рис. 3). При этом родитель, вытянув вперед голову, ожидает, пока один из них после нескольких неудачных попыток не сумеет схватить корм и проглотить его. Тогда предлагается новый кусочек и иногда еще несколько. Наконец птенцы перестают выпрашивать корм, родитель проглатывает его остатки и снова усаживается обогревать потомство.
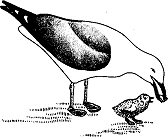
Рис. 3. Серебристая чайка кормит птенца
Другие взаимоотношения между родителями и птенцами становятся заметными, когда в колонию проникают хищники. Собаки” лисы и люди вызывают самую интенсивную реакцию чаек. Взрослые птицы издают хорошо известный крик тревоги “гага-га! га-га-га-га-га!” и взлетают. Этот крик выполняет двойную коммуникационную функцию. Птенцы бегут в укромные места и припадают к земле, а взрослые продолжают летать, готовясь к нападению. Однако настоящие атаки на пришельца совершаются индивидуально. Каждая птица устремляется вниз и может даже ударить хищника одной или обеими ногами, когда он приблизится к гнезду. Иногда атака сопровождается “бомбардировкой” Отрыгнутой пищей или фекалиями, т.е. очень неприятным оружием. Однако такие нападения не приносят полного успеха. Они лишь тревожат и отвлекают лису, собаку или человека, которые, естественно, уже не способны искать добычу так же внимательно, как в спокойном состоянии. Они не замечают какие-то гнезда, и особенно птенцов, но вполне могут наткнуться на них случайно. Впрочем, такая относительная неэффективность свойственна всем биологическим функциям: ни одна из них не ведет к абсолютному И полному успеху, но каждая способствует его достижению. Большую помощь при защите от хищника оказывают покровительственная окраска и поведение птенцов. Действительно, припадание к земле (рис. 4) скрывает их от взгляда хищника, полагающегося в основном на свое зрение.
Спустя примерно сутки с момента вылупления птенцы становятся более подвижными. Они ползают по родительской территории, постепенно удаляясь все дальше от гнезда, однако не покидают ее, пока не будут вынуждены сделать это в результате частого появления человека, например толп любителей природы. Слишком часто эта любовь становится смертельной угрозой для птенцов, поскольку, заходя на чужую территорию, они подвергаются нападению и соседи часто их убивают. Настоящий любитель природы мог бы получить больше удовольствия от терпеливого наблюдения за жизнью чаек с расстояния. Большинство из описанных выше событий заметны издалека.

Рис. 4. Затаившийся птенец серебристой чайки
Таким образом, на примере чаек можно видеть множество признаков социальной организации. Частично она служит целям спаривания. Однако некоторые формы сотрудничества между самцом и самкой не имеют с ним ничего общего и направлены на сохранение семьи. Помимо этого, наблюдается взаимодействие между родителями и потомством. Птенцы требуют от родителей корма, а те временами заставляют их прятаться и сидеть тихо. Заметно также взаимодействие между различными парами, а крик тревоги поднимает в воздух всю колонию. Результат всего этого —выращивание большого числа молодых птиц, явление настолько обычное, что упоминание о нем кажется общим местом, однако даже слабые нарушения социального поведения могут оказаться для чаек смертельно опасными. Упомянем хотя бы такой случай. Несколько раз я наблюдал, как высиживающая яйца чайка поднимается, чтобы минутку “размять ноги”. Когда она стоит и чистится метрах в двух от гнезда, другая чайка бросается сверху и клюет яйцо, разбивая его пополам. Она не успевает съесть его содержимое, поскольку родитель прогоняет разбойника, однако одно яйцо уже потеряно по неосмотрительности насиживавшего. Другой случай: в одной паре чаек самец совершенно не стремился сидеть на гнезде, не давая таким образом самке с него подняться. Она держалась героически, оставаясь на яйцах без перерыва в течение 20 суток. Однако на 21-й день оставила гнездо, и выводок погиб. Как бы ужасно это ни было для птенцов, для вида в целом такой результат благоприятен: если бы потомство унаследовало от отца описанный дефект, в стае появилось бы целых три дегенерата вместо одного.
ТРЕХИГЛАЯ КОЛЮШКА (GASTEROSTEUS ACULEATUS)
Вне периода размножения трехиглые колюшки живут косяками. Когда они совместно кормятся, можно наблюдать тип поведения, не выраженный в такой же степени у чаек, хотя он есть и у них. Если рыбе удается отыскать особенно аппетитный кусочек корма и она начинает пожирать его с типичной для колюшек жадностью, другие спешат к ней и пытаются пищу отобрать. Бывает, это им частично удается, т. е. добыча разрывается на части и делится между несколькими особями. Однако другим везет не настолько, и они начинают искать пищу на дне по соседству. Это означает, что где бы и когда бы представитель стаи ни обнаружил корм, другие стимулируются к поискам последнего тогда же и в том же месте, а это повышает вероятность обнаружения и поедания ими агрегированных видов-жертв.
Как и в случае серебристой чайки, с наступлением периода размножения формируется гораздо более сложная система социального сотрудничества, чем осенью или зимой. (Начнем с того, что самцы отделяются от косяка и выбирают гнездовые территории, приобретая при этом яркую брачную окраску. Их глаза становятся блестяще-синими, спина из мрачно-буроватой превращается в зеленоватую, а нижняя часть тела краснеет. Как только другая рыба, особенно другой самец, проникает на выбранную территорию, она тут же подвергается нападению (рис. 5). И здесь настоящие драки реже угроз, причем угрожающее поведение 
Рис. 5. Пограничная драка двух самцов трехиглой колюшки
самца колюшки весьма своеобразно. Прежде всего он бросается к противнику с поднятыми спинными иглами и открытым, готовым укусить ртом. Однако, если противник не отступает сразу, а готовится сопротивляться, хозяин территории не кусает его, а поворачивается головой вниз и, расположившись в воде вертикально, подергивается, как будто собирается закопаться рылом в песок. Часто при этом он оттопыривает один или оба брюшных шипа.
Если самца не тревожить, он начинает строить гнездо, т. е. выбрав место, зачерпывает ртом горсти песка со дна, относит их на 10—15 см в сторону и выгружает. В результате образуется мелкая ямка. Затем самец собирает гнездовой материал—обычно нити водорослей—и впрессовывает его в эту ямку. Временами он как бы ползает по этому материалу медленными подрагивающими движениями, выделяя из почек липкую слизь, склеивающую части растений. Через несколько часов или дней образуется что-то вроде зеленого клубка, в котором самец своим телом просверливает сквозной туннель.
Теперь гнездо готово. Самец сразу же меняет окраску. Красный цвет брюха усиливается, а черные пятна на спине сжимаются до крошечных точек. При этом становятся заметными искристо-голубые кристаллы гуанина, лежащие в более глубоком слое кожи, и спина приобретает яркий светло-голубой оттенок. Светлая спина при темно-красном брюхе и блестящие глаза делают внешность самца необыкновенно броской. Демонстрируя этот привлекающий наряд, он плавает вверх-вниз по гнездовой территории.
Тем временем самки, вообще не заботившиеся о гнезде, приобретают блестящую серебристую окраску, и их брюшко сильно вздувается от массивных развивающихся в яичниках икринок. Они плавают кругом косяками, иногда по нескольку раз в день пересекая выбранные самцами участки. Каждый самец, если он готов к спариванию, реагирует на самок любопытным танцем (рис. 6), состоящим из повторяющихся фигур. Сначала самец отворачивается, как будто собирается уплывать от самок, а затем резко направляется к ним с широко раскрытым ртом. Иногда он может ударить самку, но обычно останавливается прямо перед ней, а затем отворачивается, чтобы начать фигуру снова. Этот зигзагообразный танец распугивает большинство самок, но одна из них может оказаться достаточно зрелой для икрометания и, вместо того чтобы скрыться, разворачивается к самцу, одновременно принимая более или менее вертикальное положение. Тот немедленно устремляется к гнезду. Самка следует за ним. Достигнув гнезда, самец всовывает рыло в входное отверстие и ложится на бок спиной к самке, которая теперь стремится пролезть в гнездо. С помощью мощных ударов хвоста ей удается проскользнуть в узкое отверстие. При этом ее голова выступает с одного конца гнезда и хвост—с другой. Теперь самец начинает тормошить ры
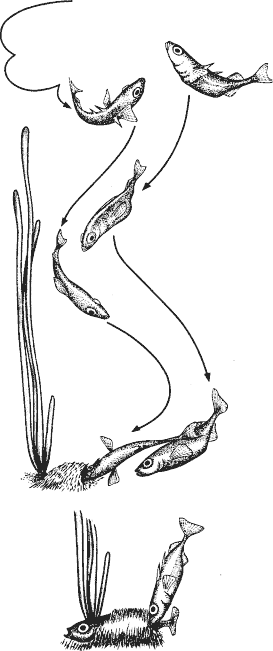
рис. 6. Последовательность действий при ухаживании у трехиглой колюшки
лом основание ее хвоста, т. е. наносит по нему серию быстрых ударов. Спустя некоторое время самка поднимает хвост и выметывает икру. После этого она спокойно выползает из гнезда, в которое заходит самец, который, затем также выскальзывая с другой его стороны, оплодотворяет икру. Теперь он отгоняет самку, возвращается к гнезду, восстанавливает его крышу, приподнятую и нарушенную в результате прохождения под ней двух партнеров, и часто перемещает и укладывает икринки так, чтобы все они оказались внутри постройки. Это конец брачных отношений. Нет ни “свадьбы”, ни продолжительной связи особей между собой, вся задача самки при размножении—выметать яйцеклетки. Забота об икринках и молоди—функция самца. Таким образом, взаимодействия между самцом и самкой сводятся к серии быстрых реакций друг на друга, которые можно представить следующей схемой:
САМЕЦ САМКА
Появление
Зигзагообразный танец
Ухаживает
Ведет самку
Следует за самцом
Показывает вход в гнездо
Входит в гнездо
Тормошит самку
Мечет икру
Оплодотворяет икру
Самец может за несколько дней “соблазнить” двух, трех и более самок, собрав в гнезде несколько кладок икры. Затем его половое влечение угасает, и ухаживание сменяется родительским поведением. Оно заключается в изгнании чужаков (рыб обоего пола своего вида, других видов, прочих хищников) и в вентиляции икры. Последнее осуществляется за счет характерного “обмахивающего” движения. Расположившись перед входом в гнездо под углом головой вниз, он гонит к нему воду чередующимися движениями вперед грудных плавников. Противодействуя обратному давлению воды на свое тело, он удерживается на месте взмахами хвоста. В результате вода движется в основном сверху и снизу, частично направляется к гнезду, а частично—назад. Такая активность контролируется сложной системой стимулов, поступающих от гнезда, его окружения и икры. Время, затрачиваемое на “обмахивание”, возрастает в течение следующих восьми суток. Сначала оно занимает около 200 с каждые полчаса, а к концу первой недели может требовать до трех четвертей суток. Это увеличение частично обусловлено возрастающим внутренним влечением, а частично—усиливающейся стимуляцией икринками, которые по мере развития потребляют все больше кислорода; возникающий недостаток его в среде активирует “обмахивание”.
Мальки выклевываются спустя семь-восемь дней, но еще сутки или около того остаются в гнезде, а затем начинают плавать вокруг него. При этом “обмахивание” гнезда самцом довольно резко прекращается, и теперь он внимательно охраняет потомство (рис. 7). Как только какой-нибудь из мальков начинает отплывать или, точнее, отбиваться от других, самец ловит его ртом и выплевывает в общую стаю. Потомство первое время слишком малоподвижно, чтобы ускользнуть от родителя. Однако рано или поздно кому-то это удается: можно наблюдать, как мальки внезапно один за другим устремляются к поверхности воды,

Рис. 7. Самец трехиглой колюшки охраняет потомство
касаются ее а затем быстро плывут назад. Самец часто все замечает, дается преследовать беглецов, но не может за ними угнаться и снова сбивает стаю вместе только после их возвращения. Это любопытное поведение мальков выполняет особую функцию, на поверхности они захватывают крошечный пузырек воздуха, проходящий затем по кишке и ее узкому боковому ответвлению в плавательный пузырь. Получив первую порцию воздуха, плавательный пузырь приобретает способность вырабатывать газ самостоятельно. Стремительное путешествие к поверхности, которое каждый малек совершает раз в жизни, должно быть очень быстрым по двум причинам: чтобы избежать возможных хищников и действующего из лучших побуждений родителя.
В течение следующих двух недель молодь становится все более и более самостоятельной и все дальше отплывает от гнезда. Стремление самца удержать ее в стае исчезает, но рыбки продолжают плавать вместе по собственной инициативе. Однако самец все еще охраняет потомство. Впрочем, постепенно он утрачивает к нему интерес, а заодно и свою блестящую окраску, спустя несколько недель покидает гнездовую территорию и ищет компанию “коллег”, тогда как молодь держится за рыб своего возраста.
Таким образом, проявления социального поведения у трехиглой колюшки и серебристой чайки во многих смыслах похожи. Тут и сотрудничество самца и самки перед оплодотворением икринок (хотя у рыбы взаимодействие полов не заходит дальше этого), и отношения между родителем и кладкой, родителем и потомством, между самими потомками. Наблюдаются и драки. Потомство определенным образом стимулирует отца, а он реагирует на это разными формами родительского поведения. Влияет ли сам отец на потомков (не считая возвращения их в стаю), заставляя тем самым держаться вблизи гнезда, еще неясно.
А теперь рассмотрим поведение насекомого. Возьмем для примера бабочку, называющуюся сатир Семела (рис. 8). Так уж получилось, что ее поведение известно мне лучше, чем какого-либо другого насекомого.
Гусеницы проводят осень и зиму на твердой сухой траве, растущей в аридных местообитаниях этого вида. В конце весны они окукливаются. В начале июля из куколок начинают вылетать бабочки. Часть своего времени они тратят на питание, т. е. сосут нектар различных цветков и пасоку деревьев, особенно тех, что повреждены личинками ивового древоточца, сверлящими живую древесину. Сатиров можно видеть группами по пять, десять и даже более особей, однако такие скопления вовсе не социальные образования, а всего лишь один из случаев одинакового привлечения различных особей внешними стимулами, в частности цветом и запахом пищи. Вскоре появляются признаки репродуктивного поведения. Самцы прекращают питаться, садятся на землю или на кору деревьев и держатся там наготове. Как только мимо пролетает другая бабочка, они взлетают и преследуют ее. Если эта особь оказалась самкой сатира Семела, готовой к спариванию,она реагирует на приближение самца опусканием на землю. Тот приземляется рядом, направляется к самке и останавливается прямо перед ней. Если она не реагирует на это взмахом крыльев (означающим субоптимальное половое влечение и часто отгоняющим самца), а остается неподвижной, он начинает элегантное ухаживание. Во-первых, делает несколько быстрых рывков крыльями вверх и вперед, а затем, держа их приподнятыми,
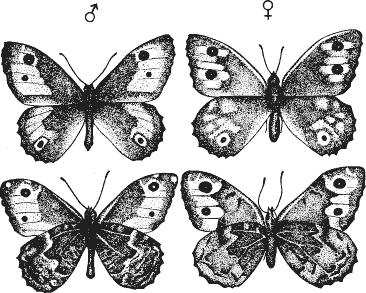
Рис. 8. Сатир Семела. Вид с дорсальной (вверху} и вентральной (внизу) сторон. Пахучие чешуйки на переднем крыле самца находятся в черном кружке
чтобы на передних было заметно красивое черное с белым центром пятно, ритмично сдвигает и раздвигает передние края крыльев, вибрируя антеннами. Это может продолжаться несколько секунд или даже целую минуту. Затем передние крылья поднимаются, широко разводятся и медленным дрожащим движением снова поднимаются так высоко, что, хотя тело самца едва движется, возникает впечатление, будто он глубоко кланяется самке (рис. 9). Затем, все еще находясь в таком положении, он сводит передние крылья, захватывая между ними антенны самки. Весь этот поклон занимает Чуть больше секунды. Затем самец разводит крылья и быстро обходит вокруг самки, оказываясь прямо позади нее. Сгибает брюшко вперед, стараясь установить контакт с копулятивными органами самки. Когда это удается, он быстро разворачивается на 180° головой от самки, и в этом положении происходит спаривание. Спустя примерно 30—45 мин самец и самка разъединяются и покидают друг друга навеки. Всю остальную жизнь эта бабочка проводит в одиночестве, никогда уже не устанавливая связей с себе подобными. Самка откладывает яйца по одному в тщательно выбранные места, где достаточно пищи для гусениц, которые не ассоциируют между собой, как у многих других видов. Таким образом, социальное поведение распространяется только на короткий процесс спаривания.

Рис. 9. “Поклон” сатира Семела
ТИПЫ СОЦИАЛЬНОГО СОТРУДНИЧЕСТВА
Сотрудничество между двумя или более особями обычно начинается с привлечения. Индивиды не просто натыкаются друг на друга, а сближаются, причем часто с большого расстояния. В апреле самцы западного соловья появляются на местах своего размножения. Их прилет легко заметить по громкому непрерывному пению. Очень приятно смотреть на них ранним утром. Часто хорошо заметно, что каждый самец ограничивает свои перемещения мелким участком земли, собственной территорией. Все самцы еще одиноки—самок пока нет. Если наблюдать за самцом изо дня в день, однажды можно обнаружить, что прилетела самка и присоединилась к нему. С этого момента они живут парой. Зная, что каждая самка появляется в одиночку и намного позже самца, пролетев огромное расстояние с места своей зимовки в Средиземноморье до наших широт, встрече соловьев различного пола остается только удивляться. Как они находят друг друга?
Другой, не менее показательный пример — бабочка сатурния. Один из южных видов этого рода, Saturnia pyri, изучался знаменитым французским энтомологом Фабром. Он сообщает, как самка, вылетевшая из куколки в неволе, окружается вскоре после этого многочисленными самцами, некоторые из которых, вероятно, пролетели для встречи значительное расстояние, поскольку вид в данной области редок. Ряд аналогичных наблюдений проведен и с другими бабочками, в частности из семейств коконопрядов, волнянок и мешочниц.
Такие примеры поражают, поскольку демонстрируют возможности, на первый взгляд значительно превышающие наши собственные. Однако в принципе они не загадочней способностей многих других видов, особи которых находят друг друга на гораздо более коротком расстоянии. Комары в своих стаях, дельфины в море, гусиные семьи на фермах и бессчетное число других примеров животных, собирающихся или держащихся вместе, так же удивительны, как и два описанных выше случая. Прежде всего неизвестно, какие сенсорные органы при этом используются. Видят ли особи друг друга? А может быть, слышат или обоняют? Или используют еще неведомые нам органы чувств? А если и ясно, какой орган применяется, почему они реагируют на обращенное к ним сообщение? Как они “узнают”, что оно значит? Короче, каков механизм сотрудничества? Далее, желательно знать, для чего служит объединение особей. В случае спаривания это ясно. Но какая польза скворцам или ласточкам собираться в огромные стаи? Или же, углубляясь в детали, какова функция “поклона” самца бабочки сатира?
Когда животные собираются вместе, возникают различные типы сотрудничества. Простейший из них можно выразить формулой “Делай, как я”. Когда одна серебристая чайка спасается от опасности, другие следуют за ней. Когда домашние куры, даже только что удовлетворив свой голод, видят, что одна из них снова начинает есть, они присоединяются к ней точно так же, как уже упоминавшиеся трехиглые колюшки. Этот принцип “симпатической индукции” (по терминологии Макдугалла) заметен в действиях многих социальных животных, включая человека. Мы зеваем, видя, как зевает другой, пугаемся, заметив признаки сильного страха у другого человека. Речь здесь не о подражании; реагирующие особи на научаются выполнять определенные движения, наблюдая за действиями других; просто у них появляется такой же настрой и происходит врожденная двигательная реакция.
Наблюдения за воздушными маневрами стаи скворцов или Других птиц дают представление о другом типе сотрудничества. Эти животные не только летают, когда летают другие, но и так же направляют свой полет. Поразительно видеть, как тысячи скворцов, кружащие зимним вечером вокруг места своего ночлега, сворачивают будто по команде. Их сотрудничество настолько совершенно, что забываешь об отдельных особях и автоматически думаешь о птицах как о едином облаке, как об одном огромном “сверхиндивиде”.
Во всех этих случаях участвующие во взаимодействии животные занимаются одним и тем же. Однако при многих других типах сотрудничества наблюдается разделение труда. Например у хищных птиц самец обычно добывает корм для всей семьи, а самка охраняет выводок. Самец приносит пищу к гнезду, но сам птенцов не кормит, а протягивает добычу партнеру (рис. 10), который и передает ее потомству. Многие птицы начинают снова размножаться, когда “первая смена” птенцов еще не способна заботиться о себе. Это значит, что родителям приходится насиживать яйца и охранять потомство одновременно. У козодоя обязанности разделены: самец находится при выводке, а самка сидит на яйцах. У кулика-галстучника партнеры периодически подменяют Друг друга: гулявший с птенцами возвращается на гнездо, а насиживавший яйца уходит к потомству. Естественно, здесь необходимы тесное сотрудничество и синхронизация действий.

Рис. 10. Самец пустельги передает добычу самке
Разделение труда достигает кульминации у медоносных пчел. Яйца здесь откладывает только матка. Задача самцов (трутней) заключается лишь в оплодотворении девственных маток. Все прочие обязанности выполняются рабочими особями, т. е. бесплодными самками. Некоторые из них строят соты, другие кормят личинок, третьи охраняют улей и отгоняют врагов, четвертые вылетают на сбор нектара и пыльцы и т.д.
Разделение труда часто требует четкой согласованности действий. Хорошие примеры тому;—брачное поведение бабочки сатира и, естественно, многих других видов. Ухаживание самца стимулирует самку к взаимодействию, а во время самого спаривания движения особей и их половых органов точно согласованы. Способы такого сотрудничества у различных видов неисчерпаемы. Часто оно еще сложнее, чем у упоминавшихся выше видов,— стоит лишь подумать о стрекозах, кальмарах, змеях или тритонах. Однако даже в простейшем случае перед исследователем встают нерешенные проблемы. Каждый, кто потратит хотя бы час времени, наблюдая за черным дроздом или другой певчей птицей, кормящей своих птенцов, увидит такой тип сотрудничества в действии. Пока родители летают за кормом, потомство лежит в гнезде спокойно. Однако, стоит взрослой птице опуститься на край гнезда, птенцы вскакивают, вытягивают шею и распахивают клюв (рис. 11). Родитель реагирует на это, наклоняясь и опуская пищу в рот одному из птенцов. Тот проглатывает полученное и успокаивается. Только это еще не конец: обычно птица чего-то ждет, пристально вглядываясь в гнездо. Скоро в куче потомства можно заметить движение: один-два птенца начинают трясти животами, кольцо игловидных перьев вокруг их клоаки расходится, и из нее появляется чистый белый комок фекалий. Он подхватывается родителем, проглатывается им или уносится и выбрасывается на некотором расстоянии от гнезда. Таким образом в результате сотрудничества осуществляется чистка последнего: птенец ловко предъявляет родителю нечистоты, а тот их устраняет.

Рис. 11. Черный дрозд кормит птенцов
У некоторых рыб можно видеть другой пример согласованного поведения. Самка крупного цихлового Tilapia natalensis захватывает икринки ртом сразу же после того, как самец их оплодотворил, и там их вынашивает. Когда выклевываются мальки, они сначала остаются во рту, но через несколько дней стайкой выплывет наружу и остаются поблизости от матери. В случае опасно-9 потомство устремляется к материнскому рту (рис. 12), она пускает его внутрь и держит там, пока тревога не минует. Число таких коротких рассказов о сотрудничестве между особями, безусловно, можно умножить. Спектр феноменов, наблюдаемых в царстве живого, гораздо разнообразнее, чем я в силах осмотреть в этой небольшой книжке.
В целом все такие примеры можно сгруппировать в четыре категории, каждая из которых иллюстрируется одним или более из осмотренных выше видов.
Во-первых, самец и самка встречаются для спаривания. Их взаимодействие приводит к оплодотворению и росту новых особей. Оно служит цели, которая не может быть достигнута каждым из партнеров в одиночку. Обычно активную роль при этом играют и самец, и самка, хотя, как правило, самец более активен.

Рис. 12. Мальки Tilapia natalensis возвращаются к матери
Во-вторых, родители (или один из них) охраняют потомство или заботятся о нем до тех пор, пока в этом есть необходимость. Здесь отношения по своему результату односторонние. Родители “помогают” молоди, а та им не “помогает”. Однако, как позволяют предполагать первые поверхностные наблюдения и подтверждает анализ, и здесь речь идет именно о сотрудничестве, поскольку потомство стимулирует родителей, вызывая их реакции, и наоборот.
В-третьих, ассоциация особей распространяется у многих видов за пределы семейной жизни на групповое сосуществование. Поскольку в последнем можно найти так много аспектов, общих с семейной жизнью (вполне вероятно, что часто это не что иное, как экстраполяция семейных отношений), семейная и групповая организации будут обсуждаться вместе.
Наконец, особи могут взаимодействовать совсем иным путем—драться. На первый взгляд драка кажется прямой противоположностью сотрудничества, т.е. антагонизмом. Однако я надеюсь показать, что схватка между особями одного вида, хотя и не приносит пользы им самим, крайне важна для вида в целом, как бы парадоксально это ни звучало. Опасность для особи здесь в принципе такая же (хотя и отличается по степени), что и при спаривании или защите потомства. Очевидно, однако, что спаривание и охрана молоди служат сохранению вида, в то время как при драке это не сразу заметно. В гл. 4 будет показано, какую функцию она выполняет и почему ее также следует считать одним из типов сотрудничества.
В следующих главах мы обсудим разнообразные способы организации видами этих четырех типов социального сотрудничества, они будут сгруппированы в зависимости от функции, которой служат, а не в соответствии с лежащими в их основе механизмами.
ГЛАВА 2
ФУНКЦИИ БРАЧНОГО ПОВЕДЕНИЯ
Многие животные, особенно морские, обеспечивают оплодотворение яйцеклеток таким простым способом, что говорить о каком-то особом брачном поведении затруднительно. Например, устрицы просто извергают в воду огромные количества своих спермиев в определенное время года, каждая особь окружается целым их облаком. По-видимому, яйцеклетки просто не могут избежать оплодотворения. Однако даже здесь различим важный поведенческий аспект: оплодотворения не произойдет, если различные особи устриц не образуют свои спермии и яйцеклетки одновременно. Значит, необходима определенная синхронизация. Я надеюсь доказать, что это относится и к наземным видам.
У многих высших животных, особенно сухопутных, оплодотворение требует спаривания, или копуляции. Для этого необходима не только синхронизация, но и физический контакт особей. Его большинство животных избегают. Избегание—адаптация, частично обеспечивающая защиту от хищников: прикосновение к телу обычно означает, что тебя поймали. Кроме того, во время самого спаривания животные (особенно самки) находятся в беспомощном положении. Другими словами, брачное поведение нередко включает в себя подавление защитных реакций. Поскольку самка некоторое время вынашивает яйца, часто даже после их оплодотворения, и поскольку у многих видов именно она играет главную роль в выкармливании и защите потомства, она составляет наиболее ценную часть “видового капитала”. Кроме того, один самец часто способен оплодотворить несколько самок, и по этой причине отдельные самцы также менее ценны биологически. Таким образом, неудивительно, что самка больше, чем самец, нуждается в “соблазнении” партнером. Вероятно, поэтому ухаживание так часто входит в задачи именно самца. Иногда приходится “соблазнять” и его, но уже по другой причине. Самцы большинства видов в сезон размножения крайне агрессивны, и пока самкам не удастся их “умиротворить”, они вместо ухаживания могут на них напасть.
Далее, помимо синхронизации, означающей координацию спаривания во времени, требуется тесная пространственная координация: самцы и самки должны друг с другом встретиться, во время спаривания привести в соприкосновение свои половые органы, и, наконец, спермий должен найти яйцеклетку. Эта ориентация в пространстве также составляет часть брачного поведения.
Наконец, очень важно избежать спаривания с представителями других видов. Поскольку гены и крайне сложные запускаемые ими ростовые процессы у каждого вида особые, скрещивание особей разных видов приведет к сочетанию несовместимых генов, а это легко может нарушить тонко сбалансированный ход развития потомства. В результате такое спаривание часто приводит к нежизнеспособным оплодотворенным яйцеклеткам, погибающим в самом начале развития. В менее тяжелых случаях гибриды выживают, но становятся слабыми или бесплодными. Необходимость избегать межвидового скрещивания привела к формированию видоспецифичных особенностей брачного поведения, так что каждая особь без труда “распознает” партнера собственного вида.
Таким образом, кроме собственно осеменения к функциям брачного поведения относятся синхронизация, ухаживание, пространственная ориентация и репродуктивная изоляция.
В этой главе мы зададимся вопросом: как выполняются перечисленные функции? Какую роль играет социальное поведение и как оно приводит к нужным результатам? Скажем прямо с самого начала, что наши знания в этой области весьма фрагментарны. По каждой из проблем собрана кое-какая информация, но частично она относится к одному виду, частично—к другому и т.д. Полного представления пока нельзя составить ни по одному из животных. Значит, единственное, что нам доступно,— рассмотреть на ряде примеров способы выполнения функций брачного поведения, оставив на будущее раздумья о том, насколько здесь возможны обобщения.
Одно очевидно сразу—обсуждаемые действия осуществляются на относительно низком “психологическом” уровне и не включают в себя ни предвидения целей, ни обдуманных действий для их достижения. Как будет показано, брачное поведение у всех видов, исключая человека и, возможно, некоторых высших обезьян, состоит из непосредственных реакций на внутренние и внешние стимулы. Здесь нет места “прогнозированию” последствий поступков, каким-то совершенно загадочным способом выступающему в роли их причины у человека.
НЕКОТОРЫЕ ПРИМЕРЫ СИНХРОНИЗАЦИИ
Хронология репродуктивного поведения у устриц, как было недавно показано, регулируется довольно неожиданным внешним фактором, поэтому, строго говоря, не относится к “социологической” проблематике. Однако не исключено, что полезно будет рассмотреть ее здесь в качестве примера того, как действие внешних факторов может, так сказать, “имитировать” социальное сотрудничество.
Примерно через восемь дней после икрометания устриц начинается “роение” их личинок. Они очень недолго плавают, а затем оседают на твердый субстрат. В илистых эстуариях Шельды в Голландии устрицеводы увеличивают свои “стада”, укладывая на морское дно черепицу, служащую личинкам устриц искусственным субстратом. Этого нельзя делать задолго до массового появления личинок, поскольку куски черепицы будут быстро заняты другими организмами. Таким образом, от зоолога требуется определить, если это возможно, когда произойдет “роение”. Прогноз, основанный на многолетних исследованиях, звучит удивительно: “Основной максимум появления личинок следует ожидать ежегодно между 26 июня и 10 июля, примерно через 10 дней после новолуния или полнолуния” (рис. 13). Это кажется мистикой, но является чистой правдой. Поскольку “роение” происходит через восемь дней после икрометания, значит, самого икрометания следует ожидать через два дня после новолуния или полнолуния. Отсюда выводится фактор, определяющий время размножения, — прилив. Икрометание происходит при сизигийном приливе. Как он влияет на устриц, еще неизвестно, не исключено, что это связано с давлением воды, колебания которого максимальны именно при сизигийном приливе. В это же время больше всего колебания освещенности на дне моря: чем не фактор?
Поскольку устрицы не мечут икру при каждом сизигийном приливе, должен быть и другой фактор, подготавливающий их именно к июньской реакции на такой прилив; природа его еще неизвестна. Он действует не слишком точно, поскольку, хотя максимум икрометания приходится на период 18 июня—2 июля, меньшие по высоте пики наблюдаются и при других сизигийных приливах. Для устриц этот фактор неизвестен, но в случае других животных мы кое-что о нем знаем.
Время размножения связано с приливами не только у устриц, но и у некоторых других обитателей моря. Среди них знаменитый тихоокеанский червь палоло и еще несколько видов червей и моллюсков.
Временной фактор у высших животных—более сложная проблема. Кое-что известно о рыбах, птицах и млекопитающих северной умеренной зоны. Размножение у большинства из них начинается весной. Первая фаза—миграция к местам размножения. Она происходит у всех особей вида примерно одновременно, хотя между прибытием на место первой и последней из них иногда проходят недели. Эта грубая синхронизация обусловлена опять-таки не социальным поведением, а реакцией на внешние факторы. В данном случае главный из них—удлинение светового дня в конце зимы. Известно, что различные млекопитающие, птицы и рыбы реагируют на искусственное удлинение долготы дня. В результате их гипофиз начинает секретировать гормон, который в свою очередь влияет на рост половых желез. Они в свою очередь секретируют половые гормоны, а действие последних на центральную нервную систему запускает первую поведенческую реакцию, связанную с размножением,—миграцию. Дополнительное влияние часто оказывает повышение температуры среды.
Как уже говорилось, эта синхронизация не очень точная. Отдельные особи реагируют на удлинение светового дня неодинаково интенсивно, причем могут наблюдаться значительные различия даже между самцом и самкой одной пары. В случае голубей и некоторых других животных было замечено, что, если самец реагирует раньше самки, его настойчивые ухаживания могут ускорить гормональное развитие последней. Это обнаружилось следующим образом. Когда самца и самку держали раздельно в соседних клетках так, чтобы они могли видеть и даже касаться друг друга, но не копулировать, непрерывное ухаживание голубя в конце концов индуцировало голубку снести яйца. Естественно, они оказались нежизнеспособными. В неволе бывает, что в отсутствие самцов пару образуют две голубки, одна из которых демонстрирует поведенческие особенности, обычно присущие самцу. Хотя их репродуктивные ритмы сначала могут быть разными, в результате обе несут яйца одновременно. Действия в отношении друг друга как-то стимулируют синхронизацию, причем не только поведенческую, но и связанную с развитием яиц в яичниках.
Возможно, этот эффект встречается и у других видов. Как полагал Дарлинг, он проявляется и при массовом ухаживании в парах птиц, размножающихся внутри колонии.
Однако необходимо дальнейшее повышение точности синхронизации. У всех копулирующих (да и у многих других) видов сотрудничество между самцом и самкой должно разворачиваться в соответствии с точным “расписанием”, и без тонко настроенной синхронизации оплодотворение невозможно. Только у очень немногих животных самец способен заставить самку спариваться с ним насильно. Значит, чаще всего какая-то форма очень точной синхронизации должна осуществляться на уровне долей секунды. Это обеспечивается своего рода сигнальной системой. В качестве примера рассмотрим спаривание трехиглой колюшки. В схеме брачного поведения (с. 20) стрелки означают не только временную последовательность событий, но и причинно-следственные связи: каждая реакция действует как сигнал, запускающий следующую реакцию у партнера. Так, зигзагообразный танец самца заставляет приближаться к нему самку, в ответ на это он ведет ее к гнезду, она реагирует, следуя за ним, и т. п. Это легко показать с помощью моделей или макетов. Когда самцу на его гнездовой территории показывают очень грубое подобие самки с полным икры брюшком (рис. 14), он приближается к нему и исполняет зигзагообразный танец. Как только модель разворачивается в его направлении и “плывет” навстречу, он начинает вести ее к гнезду.
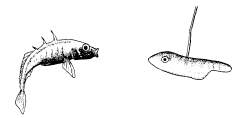
Рис. 14. Самец трехиглой колюшки ухаживает за грубым макетом самки
Самку с икрой можно аналогичным образом заставить реагировать на модель самца. И тут будет достаточно самой грубой подделки, лишь бы нижняя часть макета была красной, а глаза ярко-голубыми. Остальные детали не важны. Если такую модель перемещать вокруг самки, очень поверхностно имитируя зигзагообразный танец, самка развернется и приблизится к ней. Если теперь модель “поплывет” прочь, самка последует за ней и можно даже заставить ее попытаться проникнуть в любую дырку на дне аквариума, продемонстрировав моделью самца “вход в гнездо” (рис. 15). В самом гнезде нет необходимости, движения модели—достаточный стимул для реакции самки.
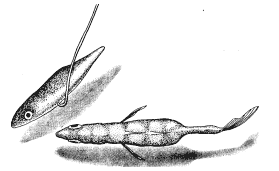
Рис. 15. Самка трехиглой колюшки следует за макетом самца, “показывающим вход в гнездо”. Вид сверху
В этих случаях рыба реагирует не только на перемещения партнера, но и на некоторые черты его формы и окраски. Если у макета самки “брюхо” не вздуто, танец самца, если и будет стимулироваться, то очень слабо. Если у модели самца нижняя часть не красная, самка не проявит к ней интереса. С другой стороны, все прочие детали практически не важны, поэтому легче запустить брачное поведение самца, используя очень грубую, но “икряную” подделку, чем живую, но не готовую к размножению самку. Однако сами по себе ни вздутое, ни красное брюхо, сколько бы их ни предъявлять, не вызывают синхронной реакции у соответствующего пола. Она возникает только в ответ на определенное движение, которое таким образом и обусловливает точную синхронизацию событий.
Брачное поведение трехиглой колюшки—это сложный ряд таких сочетаний сигнал—ответ, приводящих в конечном итоге к оплодотворению самцом икринок сразу же после их выметывания самкой. Наблюдать подобные действия и проводить описанные выше опыты с моделями совсем не трудно. Эта рыба легко размножается в аквариуме емкостью около 20 л. На дно следует насыпать песок и поместить побольше зеленой растительности, включая нитчатые зеленые водоросли.
С такими сигнальными движениями, непосредственно обеспечивающими высокую точность синхронизации, связано брачное поведение многих видов.
Даже когда животное сексуально возбуждено, оно не всегда сразу же реагирует на ухаживание партнера. Преодоление нежелания самки спариться может потребовать много времени. Так, зигзагообразный танец колюшки не всегда немедленно вызывает ее реакцию. Самка может нерешительно приблизиться к самцу и остановиться, когда он попытается увести ее к гнезду. В этом случае самец возвращается и снова исполняет зигзагообразный танец. После нескольких его повторов самка в конце концов иногда (но не всегда) соглашается следовать за ним и входить в гнездо.
Сходное повторение сигналов необходимо, когда самка уже в гнезде. Чтобы она выметала икру, самец должен ее как следует “потормошить”. Если убрать его сразу, после того как самка вошла в гнездо, икрометания не произойдет. Однако, если мягко дотрагиваться до нее стеклянной палочкой, имитируя совершаемое самцом тормошение, икра будет отметана так же легко, как и в случае стимуляции, исходящей от него самого. И самец, и палочка должны дотрагиваться до самки многократно.
У многих видов такое повторение сигналов—правило. Например, копуляции шилоклювок предшествуют любопытные ужимки: самец и самка стоят рядом и как-то торопливо, “нервно” чистят свои перья. Спустя некоторое время самка прекращает охорашиваться и вытягивается горизонтально (рис. 16). Это сигнал, что она готова к спариванию. Только теперь самец взлетает на нее, и происходит копуляция. Иногда он реагирует не сразу, а через некоторое время.
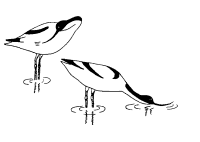
Рис. 16. Предкопуляционная демонстрация у шилоклювки
У серебристой чайки копуляции предшествует аналогичное вступление. И самец, и самка неоднократно вскидывают вверх головы, каждый раз издавая при этом мягкие, мелодичные призывы (рис. 17). В данном случае инициативу берет на себя самец: после ряда таких сигналов он неожиданно вскакивает на самку и спаривается с ней.
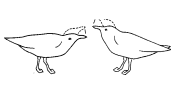
Рис. 17. Предкопуляционная демонстрация у серебристой чайки
Иногда “соблазнение” выполняет другую функцию. У многих птиц, да и у других животных самцы в сезон размножения становятся очень агрессивными. В самом деле, большинство сражений в природе происходит весной между соперничающими самцами. В этих драках вся причина. Поскольку нападения всегда направлены против самца-соперника, самка должна отличаться от него, чтобы не быть атакованной. У таких видов, как зяблик, горихвостка или фазаны, этой цели частично служат различия в оперении. Однако у многих других видов, например у крапивника, окраска обоих партнеров почти одинакова, поэтому самке, чтобы подавить агрессивность самца, приходится особым образом себя вести. Значит, задача такого “самочьего ухаживания”— постараться не спровоцировать нападения. Если самец-чужак либо спасается от демонстрирующего агрессивность хозяина территории (что немедленно индуцирует преследование), либо хорохорится и угрожает в ответ (что также вызывает демонстрацию агрессивности), самка ничего такого не делает. У рыбы горчака в ответ на первую атаку она либо спокойно отступает, либо избегает столкновения, проплывая под самцом. Видимо, после этого самец уже не способен напасть на нее, спустя некоторое время полностью утрачивает агрессивность и начинает ухаживание (рис. 18), Сходное ненавязчивое умиротворение можно наблюдать у многих цихловых рыб. В других случаях самка демонстрирует инфантильное поведение, т. е. прибегает к такому же способу умиротворения, что и потомство, возможно, стимулируя тем самым у самца родительские побуждения. Именно поэтому у очень многих видов самцы при ухаживании кормят самок.
Уже говорилось, что такое свойственно серебристой чайке. Однако у некоторых видов умиротворяющая поза при ухаживании отличается от используемой ювенильной особью. Самка (а иногда и оба партнера) демонстрирует поведение, которое во многих смыслах является противоположным угрожающему. Когда, например, обыкновенные чайки встречаются в сезон размножения, они применяют “впередсмотрящую демонстрацию”, опуская голову и направляя клюв друг на друга (вкладка 2, вверху). Эта угрожающая поза подчеркивается коричневой окраской их “лица” вокруг клюва, являющегося серьезным оружием. В то же время половые партнеры демонстрируют свои лучшие намерения с помощью, так сказать, “отмашки головой” (вкладка 2, внизу), т. е. вытягивая вверх шеи, резким рывком отворачиваются друг от друга. При этом, поскольку оба пола достаточно агрессивны, самец умиротворяет самку, а самка — самца. У некоторых тенетных пауков самец посещает самку на ее паутине. В данном случае он должен ее умиротворить, чтобы она не приняла его за добычу.
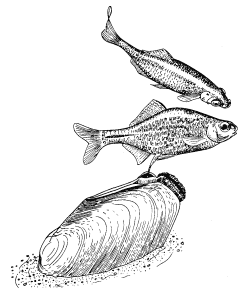
Рис. 18. Самец горчака ухаживает за самкой во время икрометания
Пространственное управление брачным поведением—еще одна важная функция ухаживания. Наиболее очевидный ее пример—привлечение. Многие певчие птицы, например соловьи, проводят зиму вдали от гнездовых участков. Самцы, как уже говорилось выше, возвращаются с юга гораздо раньше самок. Как же самки их находят? По песне. Многие птицы привлекают противоположный пол какими-нибудь громкими звуками. У соловья эти призывы приятны человеку, поэтому он называет их песней. А вот весенние крики самца серой цапли очень грубы и человеческому уху отвратительны. Однако самку этого вида они привлекают и выполняют точно такую же функцию, что и соловьиное пение. К той же категории звуков относятся треск козодоя, барабанная дробь дятла и кваканье жаб (вкладка 3, внизу). Пение многих птиц так точно настроено на привлекающую функцию, что достигает максимальной громкости, когда самец одинок, и прекращается, как только у него появляется самка. Здесь перед нами опять же столкновение различных интересов. Пение служит для привлечения самок (и, как будет показано ниже, для отпугивания самцов-соперников), но опасно для самца, поскольку выдает его хищникам. Природа, как всегда, пошла на компромисс: песня звучит до тех пор, пока действительно нужна или по крайней мере пока ее преимущества перевешивают недостатки.
Поскольку многие животные глухи (только позвоночные и некоторые другие группы составляют исключение), звуковые демонстрации встречаются относительно редко. Они хорошо развиты у птиц, жаб и лягушек, а также у ряда насекомых, включая сверчков и кузнечиков. У этих животных сформировались особые органы, предназначенные исключительно для издавания звуков.
Другие группы используют для привлечения противоположного пола запах. Крайнее выражение этой способности наблюдается у бабочек. Некоторые исследования проводились на семействе мешочниц, которое и будет рассмотрено в качестве примера. Самка здесь утратила способность к полету и практически бескрыла. Вскоре после выхода из куколки она покидает трубчатый чехол из листьев, в котором проходило ее предшествующее развитие, и подвешивается к нему снизу. Самцы могут летать. Выйдя из куколки, они оставляют чехол и отправляются на поиски самок. Их путь направляется запахом, испускаемым девственной самкой. Такой способ привлечения высоко развит и у многих других бабочек, включая виды павлиноглазок (рис. 19) и коконопрядов, причем самец часто способен отыскать самку на весьма значительном расстоянии, поскольку его органы обоняния, расположенные на перистых антеннах, высокочувствительны. Гусениц этих видов нетрудно обнаружить, собрать, дать им окуклиться и выйти из куколки, чтобы затем пронаблюдать, как в дом залетают посторонние самцы, привлеченные запахом девственных самок.
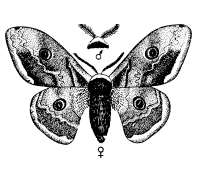
Рис. 19. Зашгта руп. У самца на антеннах сильно развиты хеморецепторные органы
У многих видов важно зрительное привлечение. Оно прекрасно развито у трехиглой колюшки. Самец приобретает особенно яркую брачную окраску, окончив строить гнездо. Его красное брюхо становится еще более насыщенным по цвету, а темный оттенок спины сменяется флуоресцирующим голубоватым. Одновременно меняется и его поведение. Если во время постройки гнезда самец движется мягко, избегая рывков, то теперь он мечется по своей территории, что вместе с броским внешним видом делает его заметным издалека.
Многие птицы дополняют звуковое привлечение зрительными демонстрациями. Это наиболее развито у видов обширных равнинных пространств. Так, в частности, специализированы многие кулики арктической тундры и европейских болот (рис. 20). Причем опять же часто наблюдается сочетание броской окраски и особых движений. Чибис, большой веретенник, чернозобик — лишь некоторые типичные примеры. У других видов специализированы только привлекающие движения, а внешность неприметная, к ним относятся наиболее уязвимые для хищников певчие птицы типа коньков и жаворонков. Встречается и чисто цветовая специализация. У самца турухтана нет особого брачного полета или призывной песни, но он может рассчитывать на окраску своего горлового оперения. Впрочем, у него развилось другое сигнальное движение: время от времени самцы на “току” поднимают крылья, светлая нижняя сторона которых делает их очень заметными (вкладка 4, вверху). Такой демонстрацией они в основном реагируют на пролетающих вдали самок; по-видимому, она их привлекает. Эти токующие птицы применяют и гак называемый “принцип клумбы”: когда они собираются вместе, их индивидуальные расцветки сочетаются друг с другом и образуется крупное пестрое пятно, напоминающее клумбу.

Рис. 20. Чибис в полете
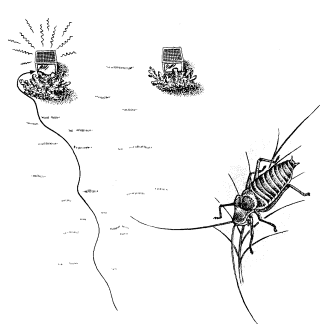
Рис. 21. Эксперимент, демонстрирующий функцию песни у кузнечи-кового Ер^рр^ег
Лишь в немногих из этих случаев привлекающее действие доказано экспериментально. Так, ясно, что самки колюшки реагируют на красный цвет брюха самца, модели с другой окраской на них не действуют. Роль звука изящно продемонстрирована для различных кузнечиковых. Один из таких опытов изображен на рис. 21. В клетке, спрятанной в зарослях вереска, держали стрекочущих самцов эфиппигера; в другой клетке находилось столько же самцов, которые не могли издавать звуков, потому что их стридуляпионные органы были склеены экспериментатором. Это простейшая операция, позволяющая таким бескрылым формам свободно осуществлять любую другую активность. На расстоянии 25 м от клеток были выпущены готовые к спариванию самки. Они безошибочно устремились к клетке со стрекочущими самцами.
Эксперименты такого типа подтверждают выводы, сделанные выше относительно привлекающего воздействия различных типов демонстраций, хотя, конечно, у экспериментаторов еще много работы.
Ориентационная функция ухаживания не ограничивается привлечением. Во время самого спаривания партнеры должны привести свои копуляционные аппараты в соприкосновение, что опять же требует направляющих действий. Это особенно хорошо заметно на примере многих насекомых, самцы которых обладают сложной системой класперов, требующих точной подгонки к соответствующим “обратным” частям полового аппарата самки. Однако и у проще устроенных животных типа птиц такая проблема тоже существует: самец не может привести свою клоаку в соприкосновение с клоакой самки, не реагируя предварительно на поступающие от последней ориентационные стимулы. Впрочем, об этих поведенческих механизмах известно слишком мало.
Межвидовая гибридизация происходит в природе исключительно редко. Это лишь отчасти объясняется неодинаковыми требованиями видов к местообитанию. Действительно, близкородственные формы, размножающиеся в отдаленных друг от друга географических областях или живущие в одной области, но размножающиеся в различных биотопах, предохранены от скрещивания пространственной изоляцией. Однако, даже если бы ее не существовало, обычным способом им не скреститься. Это обусловлено тем, что многочисленные сигналы, служащие для привлечения, соблазнения, умиротворения и синхронизации, у каждого вида совершенно своеобразны. Видоспецифична и соответствующая реакция. У каждого животного существует врожденная тенденция как подавать особые, свойственные только своему виду сигналы, так и реагировать исключительно на них. Однако в природе часто можно заметить половую реакцию на другой вид. Самцы бабочки сатира, которых я изучал несколько сезонов, начинают ухаживание, преследуя в полете партнера. Это брачное преследование вызывается не только самками сатира, но и другими бабочками, жуками, мухами, мелкими птицами, опадающими листьями и даже их собственной тенью на земле. Как же получается, что они никогда не спариваются с животными других видов? Сходные наблюдения, приводящие к такому же вопросу, можно провести над птицами, рыбами и многими другими животными.
Ответ, по-видимому, стоит искать в цепном характере действий, ведущих к образованию пар и оплодотворению. Когда самка сатира готова к спариванию, она особым образом реагирует на преследование партнера—приземляется. Все прочие виды, как правило, поступают наоборот: потревоженные преследующим их самцом, они улетают с максимально возможной скоростью, так что тот в конце концов от них отстает. Правда, близкородственные виды могут случайно отреагировать на него (вкладка 3, вверху), но никогда не наблюдалось, чтобы это привело к спариванию. У колюшек в принципе сходное поведение. Самец может среагировать на мелкого линя, привлеченного на его территорию зигзагообразным танцем. Однако для продолжения брачного поведения необходимо, чтобы партнер к нему подплыл. Даже если линь случайно так и поступит, он должен затем следовать за самцом к гнезду, вползать туда и выметывать икру до эякуляции колюшкой спермы. Другими словами, самка другого вида должна продемонстрировать правильную серию реакций на полную последовательность действий по ухаживанию, включая заключительное “тормошение”. А это настолько невероятно, что никогда не наблюдалось. Знаковых стимулов каждой отдельной реакции цепочки бывает и недостаточно для предотвращения реакции на другой вид, но, поскольку все эти реакции запускаются различными стимулами, их совокупности, как правило, хватает, чтобы межвидового спаривания не произошло. Это очевидно в случае видов с “взаимным” ухаживанием, так как здесь каждый пол демонстрирует серию специфических брачных действий. Но даже у таких видов, как бабочка сатир, у которых самка просто сидит на месте, пока самец выполняет сложный ритуал ухаживания, она последовательно стимулирует его; эксперименты показали, что каждое действие самца, описанное в гл. 1, запускается специфическим стимулом.
Такая специфика особенно важна в случае близкородственных видов. Как будет показано ниже, их поведенческие особенности, как и морфологические признаки, всегда очень сходны. Прошло слишком мало эволюционного времени для приобретения существенных различий. Однако у таких видов всегда наблюдается какое-либо четкое несовпадение брачных ритуалов, по крайней мере если пространственная (географическое или экологическое) или временная (неодинаковые сезоны размножения) изоляция не устранила в нем необходимости. Например, брачным поведением десятииглая колюшка достаточно похожа на своего трехиглого сородича. Однако у самцов этих видов развилась весьма различная брачная окраска. У десятииглой колюшки самец весной черный как смоль (рис. 22). Этот цвет так же привлекает десятииглых самок, как красный—трехиглых. Вместе с некоторыми незначительными поведенческими различиями неодинаковой окраски достаточно для того, чтобы скрещивание происходило редко.

Рис. 22. Самец десятииглой колюшки показывает самке вход в гнездо
Систематическое исследование проблемы репродуктивной изоляции проводилось на плодовых мушках. Первые результаты показывают, что попытки спаривания между разными видами прерывались на различных стадиях ухаживания, зависящих от используемых в опыте таксонов. Если такое прерывание происходит в серии наблюдений строго в определенный момент, значит, речь идет о специфической реакции, которую не может запустить партнер. Полученные на сегодняшний день результаты показывают, что в некоторых случаях правильный стимул не удается подать самцу, в других же “ошибку” делает самка.
Этот очень краткий и фрагментарный обзор, возможно, достаточен для того, чтобы продемонстрировать сложную природу поведенческих схем, обеспечивающих сотрудничество партнеров по спариванию. Было показано, что следует различать четыре типа функций, выполняемых ухаживанием. Это не означает, что каждое отдельное действие при ухаживании служит только одной из этих целей. Например, зигзагообразный танец самца колюшки, безусловно, обеспечивает синхронизацию, соблазнение, ориентацию и репродуктивную изоляцию, хотя различия в брачной окраске трехиглой и десятииглой колюшек объяснимы только с точки зрения изоляции. Известны также действия, связанные с синхронизацией и соблазнением, но не с ориентацией: например, самки бабочки сатира могут быть “настроены” и соблазнены ухаживанием одного самца, а затем спариться с другим. Это означает, что первый самец не ориентировал реакцию самки в свою сторону. Аналогичным образом у голубей настойчивое воркование самца не столько ориентирует на него голубку, сколько способствует овуляции ее гонад. У различных видов близкородственных дарви-новых вьюрков с островов Галапагос, как выяснилось, почти идентичные способы ухаживания за самками, однако межвидового скрещивания не происходит. Здесь репродуктивная изоляция обеспечивается отчасти различными экологическими нишами, а отчасти тем, что каждый партнер специфически реагирует на свойственную только его виду форму клюва, которая связана с особенностями потребляемой пищи. В данном случае ухаживание не имеет ничего общего с репродуктивной изоляцией и служит совсем другим задачам.
Во всех этих примерах ухаживание, как бы различны ни были его функции в деталях, характеризуется одной общей чертой— подачей сигналов, на которые реагирует партнер. Ниже природа и функция этих сигналов будут обсуждаться подробнее. Тогда станет ясно, что многие выводы и обобщения остаются пока гипотезами, поскольку экспериментальные данные еще слишком фрагментарны. Весьма полезно в этом смысле продолжение экспериментов с моделями.
ГЛАВА 3
ВВЕДЕНИЕ
Если в гл. 2 рассматривались взаимоотношения между двумя партнерами, направленные на достижение одной цели, сотрудничество внутри семьи сложнее, поскольку охватывает взаимоотношения не только между самцом и самкой, но и между родителями и потомством. Цели их деятельности также сложнее. Родители должны обеспечить убежище и пищу своему потомству, а также защитить его от хищников. При выполнении всех этих функций деятельность особей должна быть согласована во времени и ориентирована. Другие тенденции, которые могли бы ей помешать, требуют подавления. Так, у многих видов потомок генерирует стимулы, в принципе вызывающие у родителей пищевую реакцию. В другом случае исходящие от них стимулы способны запускать у потомка реакцию избегания. Кроме того, необходима репродуктивная изоляция, т.е. предотвращение реакции на потомка или соответственно родителя, относящегося к другому виду; такие реакции означали бы снижение эффективности поведения, а это в свою очередь — проигрыш в борьбе за существование. Далее, ситуация включает в себя новый элемент, отсутствовавший или по крайней мере не выступавший на первый план в отношениях между половыми партнерами, а именно защиту потомства от хищников. Это один из способов компенсировать беспомощность молодых особей.
Однако ни подавление посторонних побуждений, ни предупреждение межвидового сотрудничества не могут сравниться с такими сильными стрессами, как в случае брачного поведения, и, вероятно, именно поэтому семейные взаимоотношения не включают столь сложных церемоний, которые наблюдаются порой при ухаживании. Поскольку каждая церемония, т. е. использование сигналов, делает демонстрирующую их особь заметной, а следовательно, уязвимой, такие действия смогли развиться в процессе эволюции только тогда, когда их выгода перевешивала неизбежные недостатки. Другими словами, они не появились бы без крайней необходимости. Это могло бы показаться странным, если учесть изобилие сигналов, используемых различными видами, но сомнения исчезают, когда убеждаешься в жизненной важности таких сигналов. Обычно мы считаем социальное сотрудничество, например выращивание птенцов птицами-родителями, чем-то само собой разумеющимся. Однако оно выглядит так просто потому, что для нас привычно. Вместо того чтобы возмущаться, когда ненормальные родители бросают своих детей, следовало бы удивляться тому, что большинство родителей этого не делают, а стараются довести свою непростую и тяжелую работу до конца.
Сначала мы рассмотрим организацию семьи, а затем— группы, проанализировав в обоих случаях природу отношений, на которых основана эта организация.
Когда серебристой чайке случится найти на своей территории чужое яйцо до того, как у нее появилась собственная кладка, она не станет насиживать его, даже если оно лежит прямо в ее гнезде. И не потому, что птица не признает его за собственное—обычно она не способна отличать свои яйца от соседских. Дело в том, что у нее еще отсутствуют какие-то внутренние факторы, без которых реакция насиживания невозможна. Вне сезона размножение яйцо для чайки—просто пища. Незадолго перед откладкой собственных яиц и в самке, и в ее партнере происходят внутренние изменения, в результате которых их нервная система приобретает способность реагировать на стимул-ситуацию “яйцо в гнезде” насиживанием. Главный ответственный за это внутренний фактор у голубя и совы и, почти наверняка, у чайки—секретируемый гипофизом гормон пролактин. Однако способность к насиживанию определяется не только им, поскольку птица, хотя иногда и сидит в пустом гнезде, делает это редко и в любом случае недолго. Необходимы сами яйца; от них исходят зрительные и осязательные стимулы, запускающие у снесшейся птицы адекватную реакцию. Следовательно, здесь перед нами опять временная согласованность стадий: грубая гормональная синхронизация и более тонкая настройка поведения сигналами, вызывающими немедленный ответ.
Когда вылупляются птенцы, поведение родителей снова изменяется. Вступают в действие новые его программы типа выкармливания и охраны потомства. Эти программы у разных видов опять же неодинаковы, и было бы полезно обсудить побольше типов родительского поведения, встречающихся в природе. Однако рамки этой книжки нам этого не позволяют. Я могу лишь подчеркнуть необходимость в дальнейших (даже в чисто описательных) исследованиях, поскольку наши знания здесь еще очень неполны.
Переход от заботы о яйцах к заботе о потомстве также подчиняется грубой внутренней синхронизации, корректируемой более тонкой, обусловленной внешними стимулами. Так, в начале насиживания птица отрицательно реагирует на разбитое яйцо или птенца, однако к концу этого периода такие раздражители ее не тревожат, даже если появляются за несколько дней до вылупления собственного потомства. Следовательно, в ходе насиживания птица внутренне готовится к следующей фазе жизни. Природа этих внутренних изменений неизвестна. Точно установлено, что опять необходим пролактин, но, поскольку он активирует и насиживание, требуется и что-то еще.
Внешние стимулы обеспечивает птенец. В отношении некоторых птиц есть данные, что он стимулирует родителя, еще находясь в яйце. Скорее всего тот реагирует на писк, который слышен перед вылуплением.
Если собственную икру колюшки подменить более развитой, самец “признает” мальков-подкидышей и станет их охранять, причем “обмахивание” гнезда в момент выклева приемного потомства резко ослабнет. Однако полностью эта деятельность не прекратится и достигнет второго, более низкого, чем в норме, пика (рис. 23) в то время, когда положено появиться на свет его собственным малькам, если бы сохранилась исходная икра. Поскольку самец с ней уже не взаимодействует, второй пик, по всей видимости, обусловлен внутренними факторами.
Как только родители вступили в фазу заботы о потомстве, их деятельность, например кормление молоди, должна синхронизироваться с потребностями последней. С этой точки зрения лучше всего изучены опять же птицы. У многих певчих видов птенец, чтобы позволить родителю накормить себя, должен раскрыть клюв.
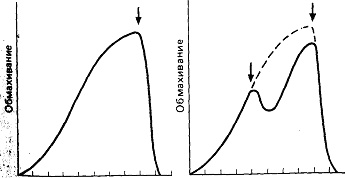
Рис. 23. Слева. Время, затрачиваемое на “обмахивание” икры самцом трехиглой колюшки в течение десяти дней после икрометания. Стрелка указывает день выклева мальков. Справа. График “обмахивания” на четвертый день собственной икры икрой, готовой к вылуплению. Вторая стрелка показывает “автономный” пик обмахивания не индуцированный чужой икрой
В противном случае птица-родитель смотрит на него, затем “беспомощно” оглядывается по сторонам, как будто недоумевает, что произошло. Она может прибегнуть к особому социальному поведению, стимулирующему потомка, например коснуться его или издать тихие призывы. Если и это не помогает, родитель обычно проглатывает принесенный корм. Природа часто дает нам возможность пронаблюдать за ее собственными экспериментами. Так, наиболее показательна реакция певчих птиц на кукушонка. Когда кукушка откладывает свое яйцо в гнездо горихвостки, птенец из него обычно вылупляется несколько раньше, чем из яиц приемных родителей. Вскоре после вылупления кукушонок выбрасывает из гнезда прочие яйца, а если за это время из них вылупляются птенцы—то и их (рис. 24). Он кладет их на спину и, пятясь назад, переваливает через край гнезда. Несчастные птенцы горихвостки гибнут от холода и голода. Они могут даже лежать на краю гнезда — все равно родители “не догадываются” вернуть их назад или обогреть и накормить там, где они находятся. Дело в том, что, поскольку кукушонок раскрывает клюв гораздо шире, родители собственных птенцов просто игнорируют—они не получают от них необходимых стимулов. В случае некоторых хищных птиц наблюдалось, что порядок кормления птенцов полностью зависит от интенсивности выпрашивания ими пищи. Самый активный в этом смысле первым получает корм. Таким образом птенцы обычно соблюдают своего рода очередь, поскольку интенсивность выпрашивания зависит от того, насколько каждый из них голоден. Если же один из них слаб с самого начала (это часто случается у луней и различных сов), не исключено, что он будет получать слишком мало пищи, в результате выпрашивать ее все слабее и в конечном итоге погибнет.
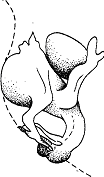
Рис. 24. Кукушонок, выбрасывающий из гнезда яйцо приемных родителей
Не только родители должны реагировать на детей, но и дети—на родителей, в частности для согласования во времени своего выпрашивающего поведения с присутствием корма. Постоянное выпрашивание, как и любой другой тип демонстрации, опасно, и такую роскошь могут себе позволить лишь немногие виды. Это было показано для некоторых селящихся в дуплах птиц, например дятлов. Но даже здесь несвоевременное выпрашивание может привести к печальным последствиям. Известно, что домовый сыч грабит гнезда таких дупловых птиц, как скворцы, и я сам видел, как ястреб одного за другим вытаскивал из дупла пищащих птенцов черного дятла. Как правило, выпрашивание ограничивается короткими интервалами появления на гнезде одного из родителей с пищей. Это обеспечивается реакцией птенцов на стимулы, исходящие от родителя. У дрозда, например, они раскрывают клювы, когда гнездо слегка встряхивается севшей на него взрослой птицей. Позднее, когда открываются глаза, они начинают реагировать на зрительные стимулы. Кроме того, потомство некоторых видов примерно через неделю после выхода из яйца способно реагировать на родительский голос. Птенец серебристой чайки стимулируется к выпрашиванию “мяукающим” призывом; в ответ на него он подбегает к родителю и начинает клевать его в конец клюва; после нескольких неудачных попыток ему удается ухватить принесенную еду и проглотить ее. Спустя еще несколько дней птенец уже различает своих родителей индивидуально и выпрашивает корм только у них. Цыплята домашней курицы более независимы и с самого начала питаются самостоятельно. Однако они сбегаются к матери, реагируя на ее особые призывы и движения, когда она находит что-либо съедобное.
Как должно быть ясно из этих нескольких примеров, функции синхронизации и ориентации обычно выполняются одними и теми же стимулами. Родители подают потомству сигнал не только типа “пора есть”, но и “пища здесь”.
Умиротворение, или подавление неадекватных реакций, ставит новые и весьма интересные проблемы. Например, у многих рыб молодь столь крохотна, что вполне сравнима с существами, употребляемыми родителями в пищу. Это относится, в частности, к колюшковым и цихловым—двум группам, у которых ярко выражена забота о потомстве. Как родителям удается не поедать собственных детей? У цихловых, “вынашивающих” мальков во рту, эта проблема решается относительно просто. Пока самка Tilapia Natalensis держит во рту потомство, она просто не питается; пищевой инстинкт полностью подавлен, а вместе с ним автоматически устраняется опасность проглотить молодь. Однако другие цихло-вые заботятся о потомстве так же, как и самец колюшки: они хватают отбившихся от стаи мальков и возвращают их назад. При этом их пищевой инстинкт не угасает, и они охотно поедают дафний, червей-трубочников и другую добычу на протяжении всего сезона размножения. Лоренц сообщает о крайне интересном и забавном случае, демонстрирующем способность родителей различать пищу и собственное потомство. Многие цихловые в сумерки уводят мальков в своего рода “спальню”, т. е. в яму, вырытую на дне. Однажды Лоренц с несколькими своими студентами наблюдал самца, собирающего потомство именно с этой целью. Только что захватив в рот одного из мальков, тот заметил необыкновенно аппетитного червячка; остановившись, самец несколько секунд смотрел на добычу и как будто колебался. Наконец, после непродолжительного “мучительного размышления” он выплюнул малька, схватил червяка и проглотил его, затем снова подобрал потомка и отнес его домой. Наблюдатели не могли удержаться от аплодисментов!
У многих птиц подросший птенец, приобретая облик взрослой особи, начинает “раздражать” своих родителей, т. е. одним своим видом стимулировать их агрессивность. Потомок может некоторое время избегать нападений с помощью инфантильного поведения, всегда однозначно понимаемого родителями. Я наблюдал это у серебристой чайки. Молодая птица принимает позу подчинения, которая в каком-то смысле противоположна угрожающей позе взрослой особи, т. е. втягивает шею, наклоняется горизонтально и слегка поднимает клюв (вкладка 5, вверху). Безусловно, неслучайно, что такая же поза принимается самкой, приближающейся к самцу, чтобы “предложить себя” (рис. 25). Однако по мере истечения периода размножения это поведение молодой птицы становится все менее эффективным, поскольку родительский инстинкт у взрослой птицы угасает. Обычно эти отношения сбалансированы так, что родители теряют интерес к потомству уже тогда, когда оно само способно следить за собой.
Другая система, не позволяющая родителям поедать собственную молодь, выработалась у цихловых рыб. Они едят мальков других видов, однако любопытный процесс научения развивает в них способность отличать свое потомство от чужого. Это было показано в простом эксперименте Нобла. Он подменил икру одной “неопытной” пары, размножавшейся впервые в жизни икрой другого вида. Рыбы признали выклевавшихся мальков и стали заботиться о них. При этом, встречая мальков собственного вида, они их съедали. Такая пара оказалась “испорченной” навсегда, поскольку в следующий раз, когда ей сохранили собственную икру, мальки были проглочены, как только из нее выклюнулись Значит, рыбы научились считать своим другой тип потомства.
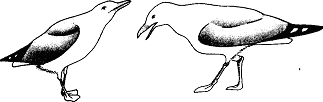
Рис. 25. Самка серебристой чайки (слева), предлагающая себя самцу
И напротив, у молоди должна быть как-то подавлена реакция на собственных родителей как на хищников. Так, мальки многих видов спасаются бегством от рыб размером с их родителей. Очевидно, это сделало бы невозможной заботу о потомстве, если бы избегание самих родителей чем-то не предотвращалось. В случае колюшки у меня сложилось впечатление, что самец просто слишком быстро плавает и успевает перехватить мальков, когда они пробуют от него ускользнуть. Забавно наблюдать за таким папашей, пытающимся удержать свое потомство в куче, когда оно изо всех сил старается разбежаться. Как уже говорилось в гл. 1, мальки, стремящиеся к поверхности для заполнения своего плавательного пузыря воздухом, развивают поразительную скорость, специально приберегаемую для этого действия. В результате они попросту обгоняют отца и успевают подняться наверх и вернуться назад, не оставляя ему шансов поймать беглецов, хотя он все время пытается это сделать. Мальки цихловых полностью доверяют своим родителям; чем это объясняется, я не знаю.
Лоренц обнаружил, как кваквам удается “убедить” своих птенцов в своих лучших намерениях. Когда родитель подходит к гнезду, он отвешивает глубокий поклон тому, кто в нем находится, неважно—партнер это или потомок. Кланяясь, птица демонстрирует свою красивую иссиня-черную “шапочку” и поднимает три тонких белых пера, растущих на голове, которые обычно сложены сзади (рис. 26). После такого вступления она заходит в гнездо, где встречает сердечный прием. Однажды, когда Лоренц, взобравшись на дерево, затаился около гнезда квакв, вернулся один из родителей. Привыкшая к наблюдениям ученого птица не обратилась в бегство, но, вместо того чтобы совершить умиротворяющую церемонию, приняла угрожающую позу. Птенец, который не боялся Лоренца, сразу же атаковал своего отца. Это было первое подтвердившееся впоследствии указание на то, что птенец “узнает” своих родителей по свойственному только кваквам (действительно, уникальному среди птиц) умиротворяющему поведению, которое подавляет его защитную реакцию.
О проблеме подавления неадекватных реакций известно немного, однако несколько этих примеров показывают, насколько она интересна. Нетрудно провести некоторые исследования в этом смысле даже в полевых условиях. С учетом рассматриваемой функции становятся понятными многие типы поведения, казавшиеся прежде бессмысленными. Например, у цихловой рыбы родители охраняют потомство по очереди. Рыба, “сменяемая” со своего поста партнером, стремглав бросается прочь от мальков (рис. 27). Берендсы показали, что это не позволяет потомству уплывать вслед за освобождающимся отобязанностей родителем.
Интересно наблюдать, как люди уже на разумном уровне разработали аналогичные церемонии. “Приветствие”, каков бы ни был его психологический подтекст, часто выполняет функцию умиротворения, подавления агрессивности и других нежелательных реакций, открывая таким образом путь для дальнейшего сотрудничества.

Рис. 26. Кваква в покое (вверху) и во время “умиротворяющей церемонии”

Рис. 27. Нет1с1иот15 Ытаси1а1ш. Освобожденный от своих обязанностей родитель быстро уплывает от потомства по прямой, что подавляет реакцию следования за ним у мальков
Функция репродуктивной изоляции, вероятно, гораздо менее важна в сфере отношений между родителями и потомством, чем в случае спаривания. Естественно, она необходима, чтобы родительская забота не распространялась на отпрысков других видов, поскольку это могло бы нанести ущерб собственным наследникам. Поведение взрослых птиц обычно обусловлено местом, где находится их потомство. В случае видов, у которых оно передвигается вместе с родителями, например у куриных, функция репродуктивной изоляции, по-видимому, важнее; весьма вероятно, что ее выполнению способствует видоспецифичный узор на голове у цыплят этой группы птиц. Звуковые сигналы у многих видов, возможно, также помогают родителям сосредоточить внимание на потомстве собственного вида.
Потребность в защите беспомощного потомства связана с новой категорией социального поведения. У многих видов молодь “замаскирована”. Однако маскировка помогает только тогда, когда сочетается с неподвижностью. Поиск пищи и ее выпрашивание, напротив, требуют движения. В результате у многих видов выработалось особое поведение, которым родители стимулируют потомство “затаиться”. Так, крик тревоги черного дрозда подавляет у птенцов выпрашивающее поведение и заставляет их застыть в гнезде. Насколько сильна эта реакция, мне пришлось убедиться, когда я утомительно долго пытался различными стимулами снова запустить у них выпрашивание. Как только родители подавали сигнал тревоги в ответ на наше присутствие около гнезда, птенцы становились безразличными к самым соблазнительным кусочкам пищи. Эта реакция еще сильнее развита у птиц, птенцы которых бродят вокруг гнезда и обычно обладают покровительственной окраской. Самые юные серебристые чайки при крике тревоги просто затихают в гнезде. Подрастая, они учатся распознавать особые убежища в окружающих зарослях, по тревоге бежать в укрытие и затаиваться в нем. У других видов потомство полагается не столько на маскировку, сколько на защиту родителей. Молодь таких животных ищет убежище около взрослого животного, заслонившись им от хищника, т.е. как бы скрывшись в создаваемой им “мертвой зоне”. Это можно наблюдать у многих уток, гусей, а также у некоторых сов. Аналогичная реакция (естественно, совершенно независимо) выработалась у цихловых.
Пример серебристой чайки—хорошая иллюстрация необходимости различать в крике тревоги синхронизирующую и ориентирующую функции. Этот крик, издаваемый родителями, определяет время реакции, заставляя птенца искать убежище. Однако родитель не способен указать, ни где находится хищник, ни куда бежать прятаться. Это стало для меня ясно когда я фотографировал из укрытия колонию серебристых чаек. Мое укрытие стояло на одном месте так много дней, что воспринималось и взрослыми и молодыми птицами как часть ландшафта. Родители использовали его крышу как наблюдательный пост, а птенцы прятались в нем при опасности. Однажды, сидя в убежище, я сделал неосторожное движение, замеченное одной из взрослых птиц, которая тут же стала тревожно кричать и устремилась от меня прочь. Потомство, взбудораженное ее криком, бросилось искать убежище. По привычке птенцы вбежали прямо в “логово льва” и затаились у моих ног.
Помимо отношений между родителями и потомством существуют и отношения между самцом и самкой, позволяющие им делить обязанности. Например, у чибиса самка сидит на яйцах, в то время как самец охраняет гнездо. Его задача—атаковать хищника и предупреждать самку о его приближении. Услышав предупреждающий крик, самка отбегает от яиц, которые остаются защищенными только покровительственной окраской. Отбежав примерно на 50 м, она взлетает и часто присоединяется к самцу, нападающему на хищника. У других видов яйца насиживают самка и самец по очереди. Здесь опять же встает проблема временной согласованности. У многих видов яйца буквально ни на секунду не оставляются родителями. Что же мешает насиживающей птице бросить их до появления своего партнера? Все дело в “освобождающей” церемонии, которой она должна дождаться и без которой ей очень трудно покинуть свое место. Когда ненасиживающий партнер в паре серебристых чаек часами ищет пищу, у него растет потребность сесть на яйца и вернуться на свою территорию. Опустившись на землю, он собирает немного гнездового материала и идет к гнезду, часто издавая при этом “мяукающий” призыв, тот же самый, что и перед кормлением птенцов. Именно такое приближение вместе с характерным криком и поведением стимулирует сидящего на яйцах партнера подняться на ноги. Однако, если его потребность насиживать еще слишком сильна, он может и не отреагировать на самую настойчивую “освобождающую” церемонию и остаться в гнезде. При этом партнер иногда пытается вытолкнуть его силой и между ними возможна спокойная, но упорная борьба (вкладка 4, внизу). У некоторых видов случаются прямо противоположные ситуации. Когда пара куликов галстучников потревожена у своего гнезда, бывает, что, даже после того как хищник удалился, обе птицы не проявляют желания вернуться к насиживанию. В таких случаях можно видеть, как самец подгоняет самку к гнезду.
Еще более сложная ситуация возникает, когда два последовательных цикла размножения “перекрываются”, т. е. когда новая кладка появляется раньше, чем предыдущий выводок стал самостоятельным. Это регулярно наблюдается у козодоя и часто у галстучника. У козодоя обязанности родителей четко разделяются: самец остается с птенцами, а самка сидит на новых яйцах. У галстучника оба родителя по очереди занимаются птенцами и сидят на яйцах (рис. 28), каждые несколько часов меняясь обязанностями. Временная координация здесь также обеспечивается особым типом поведения, сигнализирующим партнеру о необходимости смены.

Рис. 28. Галстучник с птенцами
Крики тревоги, которыми родители предупреждают друг друга об опасности, одновременно действуют и на их потомство. Однако реакции партнеров на них различны. У некоторых видов самка затаивается на гнезде. Это характерно для птиц, гнездящихся на открытом пространстве, когда самка отличается покровительственной окраской, например у многих уток, козодоев, кроншнепов и фазанов. Другие виды ведут себя как серебристые чайки и, оставляя яйца и птенцов, нападают на хищника. Такая атака может предприниматься одной парой или представлять собой настоящее коллективное взаимодействие. Например, члены колонии галок часто нападают на противника все вместе. Каждая птица не только реагирует на крик тревоги других, но и присоединяется к прочим, атакуя хищника, даже если он не угрожает непосредственно ее потомству. В случае обыкновенной крачки я наблюдал индивидуальные нападения на приближающегося человека и общие—на горностая.
У многих птиц крайне важно, чтобы оба партнера знали местонахождение гнезда, и известно множество типов поведения, демонстрирующих, где оно расположено. Когда самец и самка выбирают место для откладки яиц вместе, они с помощью особых церемоний приступают к его освоению. Так, серебристые чайки по очереди садятся на выбранное место и выкапывают лапами гнездовую ямку. У многих дупловых птиц самец, первым прибыв на подходящую территорию, выбирает дупло и привлекает к нему внимание самки с помощью особых демонстраций. Например, У горихвостки он указывает, где в него вход (рис. 29), используя Для этого ярко окрашенную голову и красный хвост. Самец пустельги планирует к гнезду особым церемониальным способом на глазах у самки.

Рис. 29. Два способа, которыми самец горихвостки указывает самке гнездовое дупло: А—демонстрирует ярко окрашенную голову; Б— демонстрирует красный хвост
Необычный случай представляет собой круглоносыи плавунчик. Здесь более броско окрашена самка, а у самца тусклое покровительственное оперение. Самка выбирает территорию охраняет ее и привлекает пением самца, а тот в одиночку высиживает и выращивает птенцов. Поскольку самка все же настолько женственна, что несется, ей приходится указывать местонахождение гнезда партнеру. Для этого она возобновляет пение непосредственно перед откладкой яйца. Привлекающее действие ее голоса не слабее, чем при первой встрече партнеров, и самец тут же устремляется на звук. Самка направляется к гнезду и откладывает яйцо в его присутствии. Это и есть церемония, приводящая самца к месту, где находятся объекты его будущей заботы.
Многие животные объединяются в более крупные, чем семьи, группы. Иногда они состоят из нескольких семей, как в случае гусиных и лебединых стай, или же образованы особями, уже не связанными семейными узами. Выгоды для индивидов от группового существования различны. Среди них наиболее очевидна защита от хищников. Высшие животные предупреждают друг друга об опасности, в результате чего группа в целом бдительна, не менее самой наблюдательной из составляющих ее особей. Более того, многие животные объединяются для совместного нападения. Эти функции наблюдаются в основном среди теплокровных, но и на более низких уровнях эволюции можно обнаружить множество других выгод групповых действий. Алли и его сотрудники экспериментально продемонстрировали много случаев таких социальных преимуществ. Например, серебряный карась в группе ест больше, чем в изоляции, и быстрее растет, причем не только из-за увеличения количества съедаемой пищи, но и под действием каких-то других факторов. Рост ускоряется и тогда, когда каждая изолированная рыба потребляет столько же корма, сколько и живущая в группе. Морской плоский червь в группе лучше переносит колебания солености, чем в изоляции. Успехи тараканов в тестах на ориентирование лучше, если их содержать по две-три особи вместе, а не поодиночке. Преимущества скоплений для дафнии, как показал Уэлти, заключается в снижении их уязвимости для хищников; когда карась сталкивается с очень плотной массой этих рачков, у него как бы “разбегаются глаза”, и он некоторое время выбирает, с какой жертвы начать. В результате общее количество добычи оказывается меньше, чем было бы при питании группой умеренной численности. Скопление гусениц бабочки Уапезш ю защищает их от певчих птиц типа горихвосток; регулярно наблюдалось, что они не нападают на группу гусениц, зато склевывают любую из тех, кто от нее отбивается.
Таким образом, очевидно, что групповое существование дает особям, а следовательно, и видам преимущества. Здесь опять же уместно спросить: как поведение способствует достижению этого благоприятного результата?
Прежде всего особи должны собраться и остаться вместе. Это может быть обеспечено сигналами, воздействующими на различные сенсорные органы. У птиц такие сигналы обычно зрительные, слуховые или и те и другие одновременно. Показано, что зеркальца на крыльях уток и гусей, выделяющиеся яркой окраской и неодинаковые у разных видов, служат именно для этой цели. Хайнрот обнаружил в Берлинском зоопарке, где вместе содержатся утиные из разных частей света, что утки и гуси, которые часто реагируют на летящую птицу, взлетая и присоединяясь к ней, наиболее охотно делают это, когда ее зеркальце напоминает их собственное независимо от таксономической близости. Яркий и своеобразный узор на хвосте многих птиц, особенно куликов, безусловно, служит для той же цели. Призывные звуки, издаваемые певчими пернатыми, например синицами и вьюрками, способствуют сохранению группы; каждая особь привлекается песней, свойственной только собственному виду, что легко заметить, наблюдая за поведением отбившейся от стаи птицы.
Многие рыбы реагируют друг на друга прежде всего зрительно, но в некоторых случаях важна и хеморецепция. Например, гольяны различают “запах” своего вида. Они даже могут научиться распознавать по нему отдельных особей, но имеет ли это значение в природных условиях, неизвестно.
Социальное поведение высших животных выходит за. рамки простой агрегации. У некоторых видов известно более сложное сотрудничество. Как уже говорилось в гл. 1, колюшки реагируют "на вид своего питающегося сородича стремлением найти пищу и для себя. Этот эффект известен под названием “симпатической индукции”, или “социального облегчения”; он наблюдался у многих видов и связан не только с питанием, но и с другими инстинктами. Когда одна птица в стае проявляет признаки тревоги, прочие также настораживаются. Сон – еще один “заразный” тип поведения. Таким способом синхронизируются даже прогулки и полет; когда несколько членов стаи демонстрируют стремление отойти прочь, прочие могут к ним присоединиться. Внезапный взлет немедленно поднимает в воздух всю стаю. Преимущества всех этих типов социального облегчения очевидны: оно синхронизирует деятельность особей, препятствуя их рассеиванию в ходе выполнения различных функций.
Большинство таких взаимоотношений основано на высокоразвитой тенденции каждого индивида реагировать на. движения других. Социальные животные чувствительны к мельчайшим знакам, к самому незначительному “шевелению”. Слабо выраженные движения типа нерешительных попыток шагнуть или подпрыгнуть называются движениями намерения, или интенционными. Многие социальные сигналы, иногда высокоспециализиро-ванные, явно относятся к их производным. Взлетая, галка внимательно смотрит на других членов группы. Если они остаются на месте, она либо возвращается назад и откладывает свое намерение до лучших времен, либо побуждает остальных присоединиться. Для этого она подлетает к сидящим на земле особям и низко планирует над ними, быстро тряся на лету хвостом.
Другой тип социального сотрудничества—совместное нападение. Оно опять же лучше всего известно для птиц. Многйе^из них, например галки, крачки и различные певчие виды, “осаждают” хищника. Они, например, могут собираться вокруг сидящего в кустах ястреба-перепелятника, или домового сыча, или же над крадущейся кошкой. Такое поведение часто можно наблюдать у домовых воробьев. В других случаях они летят плотной стаей над ястребом, стараясь держаться повыше, и время от времени на него пикируют (рис. 30).
Бывает, что такое поведение запускается у всех особей одновременно, поскольку каждая из них сразу же видит хищника. Если же он замечен только одним членом стаи, тот поднимает тревогу и предупреждает об опасности прочих. Крик тревоги—яркий пример действия, служащего интересам всей группы, но угрожающей отдельному индивиду.
У совместных нападений различные функции. Если хищник не слишком голоден, можно часто видеть, как он спешит восвояси, едва начинается такая атака. Если же ястреб голоден по-настоящему, т.е. когда он интенсивно охотится, осаждающая стая птиц его особенно не беспокоит. Тем не менее его внимание, направленное на поиск другой добычи, частично рассеивается. Даже сбивание в стаю без всякого осаждения врага, как у скворцов или куликов, преследуемых соколом-сапсаном, представляет ценность с точки зрения выживания; пикирующий на добычу сапсан старается выбрать особь, отбившуюся от прочих, поскольку из-за своей огромной скорости, пролетая сквозь густое скопление птиц, он вполне может пострадать сам.
Сигнал тревоги не обязательно должен быть зрительным или звуковым. Известно, что у многих социальных рыб его природа химическая. Когда щука или окунь выхватывают из косяка одного гольяна, другие рассеиваются и к этому месту уже не возвращаются. Долгое время они остаются начеку и бросаются в укрытие при малейших признаках появления хищника. Это обусловлено их ольфакторной реакцией на вещество, высвободившееся из кожи убитого сородича. Ее можно наблюдать и у гольяна в аквариуме, подмешав ему в корм экстракт из такой свеженарезанной кожи. Вещество это видоспецифично, как и реакция на него, причем рыбы реагируют только на “вещество страха” собственного вида.
ГЛАВА 4
Когда животное загнано хищником в угол, оно часто вступает с ним в драку. Однако этот тип борьбы—защита от хищника— нас интересовать не будет, поскольку обычно в ней участвуют особи разных видов. Она и не так распространена, как внутривидовые схватки. Большинство их происходит в сезон размножения, поэтому они называются брачными сражениями. Однако стычками сопровождается и установление иерархических отношений в группе, не связанное с определенным сезоном.
Разные виды дерутся по-разному. Прежде всего неодинаково используемое оружие. Собаки, чайки и различные рыбы друг друга кусают. Исключительно для этой цели у самца лосося в сезон размножения развиваются впечатляющие челюсти. Лошади и многие другие копытные стараются лягнуть друг друга передними конечностями. Олени меряются силой, сцепившись рогами. Во многих парках всю весну можно наблюдать, как дерутся лысухи. Они полуопрокидываются на спину и бьют друг друга своими длиннопалыми ногами. Драки многих рыб заключаются в направлении на противника сильного потока воды мощными боковыми ударами хвоста. Хотя они фактически друг до друга не дотрагиваются, движение воды существенно воздействует на их высокочувствительные органы боковой линии (рис. 32). У самцов горчака весной на голове развиваются роговые выросты, и они стараются боднуть друг друга.

Рис. 31. Драка оленей
 |
 |
Вторая важная деталь: хотя на протяжении всей весны происходит огромное число драк, относительно редко удается заметить двух животных, вступивших в насюящий “смертельный бой” и калечащих друг друга. Большинство сражений представляют собой своего рода “блеф”, простую угрозу. Причем ее действие во многом такое же, что и у настоящей схватки: попытки сближения соперников взаимно пресекаются. Некоторые примеры угрожающих демонстраций приведены в гл. 1; их разнообразие практически бесконечно. Большие синицы, стоя друг напротив друга, вытягивают вверх шеи и медленно раскачиваются из стороны в сторону, демонстрируя таким образом бело-черный узор на голове. За-рянка демонстрирует сопернику красную грудь, медленно поворачиваясь перед ним вправо и влево (рис. 33). Некоторые цихловые в качестве угрозы демонстрируют оттопыренные жаберные крышки. У СусМазота тее1с1 и Нет1сНгот15 Ытаси1а1и8 они украшены хорошо заметными черными пятнами, окаймленными золотым кольцом, и угрожающая демонстрация выглядит у них очень красиво (рис. 34).
Не у всякой угрозы зрительная природа. Многие млекопитающие оставляют “пахучие сигналы” в тех местах, где они встречают или ожидают встретить соперника. Собаки с этой целью мочатся, гиены, куницы, серны, антилопы и многие другие виды используют специальные железы, секретами которых метят почву, кустарники, пни, камни и т. д. (рис. 35). Бурый медведь чешется спиной о дерево, одновременно мочась.
Рис. 34. фронтальная угрожающая демонстрация
Рис. 35. Самец Ап1е1оре сегукарга метит дерево секретом пахучей железы, расположенной перед глазом
Звуки также могут выполнять угрожающую функцию. Все крики, упоминавшиеся в гл. 2 в качестве социально значимых “песен”, не только привлекают самок, но и служат для отпугивания самцов-соперников.
Брачными сражениями всегда занимается определенная категория особей. В большинстве случаев дерутся самцы, нападающие исключительно или главным образом на других самцов своего вида. Иногда дерутся и самец, и самка, причем, когда это происходит, самец нападает на другого самца, а самка — на другую самку. У кулика плавунчика и некоторых других видов птиц дерутся самки, причем опять же главным образом с другими самками. Все это показывает, что цель нападения—половой соперник.
Далее, драка, как и угроза, как правило, не дает двум соперникам или конкурентам обосноваться на одном и том же месте взаимная враждебность заставляет их расходиться подальше, обеспечивая себе таким образом определенное резервное пространство. Анализ того, что за смысл в этой “нейтральной полосе”, поможет нам понять значение брачных сражений.
Драка каждой особи обычно происходит на некой ограниченной площади. Это может быть, например, пространство вокруг самки (или самок), как у оленя и многих других животных. Самец горчака охраняет от других самцов участок вокруг раковины пресноводной жемчужницы (рис. 36). К этому моллюску он привлекает самку и стимулирует ее откладывать икринки в его мантийную полость, где они будут развиваться, ведя паразитическое существование. Жук-могильщик рода отгоняет соперников от падали.
Рис. 36. Самец горчака с раковиной двустворчатого моллюска
Следует отметить, что во всех этих случаях охрана относится не только к некому центральному объекту, но и к определенной площади вокруг него, т. е. соперники удерживаются на значительном расстоянии. У некоторых видов такой объект легко заметить: голубь преследует голубку и дерется только вблизи нее. Если передвинуть раковину жемчужницы, горчак переместит следом охраняемую территорию. Однако у большинства видов такая территория неизменна; самец поселяется на выбранном участке и защищает его. Территориальные стычки и угрозы можно наблюдать в любом саду: зарянки, зяблики (рис. 37) и крапивники—заядлые драчуны. Понять значение индивидуальной территории удается, если драки приурочены к какой-то ее точке. Так, у многих дупловых птиц стычки особенно яростны, когда нарушитель границы оказывается вблизи дупла. Однако у многих видов

Рис. 37. Дерущиеся самцы зяблика
драки затеваются где угодно, и тогда их значение понять труднее. Предполагают, что гнездовой участок многих певчих птиц служит как бы складом корма для потомства. Его размер должен позволять родителям собирать основное количество пищи вблизи от гнезда, т. е. сокращает расстояние, пролетаемое в ее поисках. Поскольку только что вылупившихся птенцов надо постоянно обогревать и кормить, такая территория способствует уменьшению интервалов между их посещениями. Продолжительность отлучки в неблагоприятные для птиц дни может оказаться решающей для выживания потомства. Впрочем, мнения по поводу правильности такого толкования расходятся.
У птиц, гнездящихся на земле, например чаек, крачек, чибисов и т.д., свободное пространство вокруг гнезда, по-видимому, способствует защите выводка от хищника. Ряд данных говорит о том, что слишком высокая концентрация в пространстве добычи типа яиц или птенцов заставляет хищника на ней специализироваться; это—главная причина, как правило, одиночного образа жизни и обширных гнездовых участков животных с покровительственной окраской. В случае птиц типа чаек, у которых такая окраска свойственна птенцам, территориальные драки удерживают индивидуальные выводки на достаточно большом расстоянии друг от друга. Здесь столкновение двух интересов опять же решается компромиссом: социальное гнездование дает определенные преимущества (см. гл. 3), значительные пространства между гнездами—тоже. Различные виды чаек и крачек получают в результате некоторую, хотя и не полную выгоду, следуя обеим тенденциям.
Итак, понятно, что брачные сражения выполняют определенную функцию. Они ведут к пространственному разрежению популяций, обеспечивая каждой особи обладание определенным объектом или территорией, которые необходимы для воспроизведения вида. В результате предупреждается совместное пользование такими объектами, которое во многих случаях было бы губительным или по крайней мере менее эффективным. Слишком много икринок горчака в жемчужнице сократит рацион каждой из них.
Спаривание многих самцов с одной самкой означало бы вместо создания размножающихся пар растрату сперматозоидов. Присутствие двух семей скворцов в одном дупле может привести к гибели обоих выводков. Пространственное разделение особей позволяет им полнее использовать имеющиеся возможности для выживания.
Встает второй вопрос: что обеспечивает выполнение в ходе драк названных выше функций? Другими словами, что заставляет животных вступать в бой только тогда, когда это необходимо, и только там, где надо? Как они выбирают потенциального соперника среди множества встречающихся особей? Поскольку драка для индивида опасна (она повышает его уязвимость для нападения хищника) и может поставить под угрозу успех воспроизведения (постоянные сражения оставили бы мало времени на все остальное), ограничение агрессивности только ситуациями, в которых она выполняет полезные функции, приобретает жизненно важное значение. Эти проблемы близки к обсуждавшимся в главе о спаривании. Чтобы ограничить драки именно охраной территории, раковины, самки и т. п., животное должно специфически реагировать на определенные ситуации. Кроме того, атаки должны быть синхронизированы во времени с теми моментами, когда наличествует индивид, которого надо изгнать. Наконец, усилия не должны тратиться на другие виды, за исключением тех случаев, когда они также попадают в категорию соперников. Как будет показано, многие внешние стимулы, ответственные за различные аспекты такой координации, исходят от других особей. Более того, поскольку большинство этих стимулов выполняет сразу несколько функций, я не буду останавливаться на каждой из них отдельно, как в главе о брачном поведении.
Как уже говорилось, ограниченность определенным участком—одна из самых характерных особенностей драк. Когда самец колюшки встречает весной другого самца, между ними обязательно возникает противостояние. Зависит ли его исход от места встречи? Когда1самец находится на своей территории, он нападает на всех заплывающих на нее соперников. Оказываясь за ее пределами, он обращается в бегство от того же самого самца, которого атаковал “дома”. Это можно изящно продемонстрировать в аквариуме, если он достаточно велик, чтобы заключать две индивидуальные территории. Самец а нападает на самца б, когда тот заплывает на территорию А, и наоборот. Обычно ни один из самцов не попадает добровольно на чужую территорию, но можно спровоцировать конфликтную ситуацию, отловив их и посадив порознь в стеклянные пробирки. Когда обе пробирки опущены в воду на территории А, самец а попытается атаковать самца б, несмотря на то что они разделены двойной стеклянной стенкой. При этом самец б явно постарается убежать. Когда обе пробирки перенесены на территорию Б, ситуация становится прямо противоположной (рис. 38).
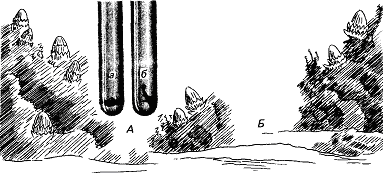
Рис. 38. А. Опыт, демонстрирующий зависимость атаки от территории. Самец б, хозяин территории Б, переносится в пробирке на территорию А. принадлежащую самцу а; последний пытается атаковать, самец б—бежать
Рис. 38. Б. Те же самцы на территории Б: самец б атакует, самец а спасается
Как своя территория стимулирует самца драться, редко учалось в деталях. Естественно, это можно сделать, только переместив территорию или ее часть и пронаблюдав, приспосабливает ли самец свою агрессивность к изменившейся ситуации. Конечно, в случае птиц такая процедура сложна из-за крупных размеров гнездовых участков, но мелкие аквариумные рыбы предоставляют в этом смысле уникальные возможности исследователю. Впрочем, сообщалось о нескольких случаях расширения птицами своей территории за пределы, изначально установленные самцом, после того, как самка начинала строить гнездо.
Для меня несомненно, что территории выбираются главным образом на основе особенностей, вызывающих врожденную реакцию животных. Это заставляет всех особей одного и того же вида или по крайней мере одной и той же популяции занимать одинаковый общий тип местообитания. Однако индивидуальная связь самца со своей собственной территорией—частным проявлением потребностей его вида—результат научения. Самец колюшки рождается с общим стремлением выбирать мелководные местообитания с обильной растительностью, но генетически заложенной тенденции реагировать на определенное расположение растений и камешков на дне у него нет. Он сдвигает свою территорию, если переместить характерные для нее “вешки”, поскольку на них у него выработался условный рефлекс. Это ясно хотя бы из того, что, размножаясь два или три раза подряд, он часто всякий раз сменяет территорию. В каждом случае рыба ориентируется по новым вешкам.
Животные, реагирующие на особые объекты типа дупла или, как горчак, на раковину жемчужницы, вероятно, делают это врожденно, т. е. для них существует всего несколько “знаковых стимулов”, связанных с этими объектами. Например, горчак ориентируется не столько на внешний вид моллюска, сколько на выходящий из его раковины поток воды. Рыба воспринимает как ее движение, так и химический состав (рис. 39).
Поступающие от собственного участка стимулы, на которые животное реагирует либо врожденно, либо в сочетании с приобретенными условными рефлексами, заставляют его ограничить площадь драк именно данной территорией.
Грубый временной контроль агрессивности также связан с внешними факторами. Как и при спаривании, первый, очень общий настрой на драку обеспечивается половыми гормонами. Воинственность отражает рост гонад, который в свою очередь через гипофиз зависит от таких ритмических факторов, как, например, долгота дня в случае многих животных северной умеренной зоны. Однако более тонкая временная настройка опять же связана с реакцией на внешние сигналы. Исходя от соперника, они вызывают нападение, когда он приближается на некоторое критическое расстояние к территории или какому-либо охраняемому объекту. У этих сигналов всегда любопытная двойственная функция. Если их демонстрирует чужак, они вызывают нападение на него, а если атакующий на собственной территории—пугают пришельца. В экспериментах с макетами можно вызвать оба типа ответа в зависимости от места предъявления раздражителя—на территории особи или за ее пределами. В обоих случаях они служат увеличению расстояния между индивидами, а, поскольку запускаются специфическими демонстрациями, а не угрозой со стороны других видов, враждебность обычно вызывают только сородичи.
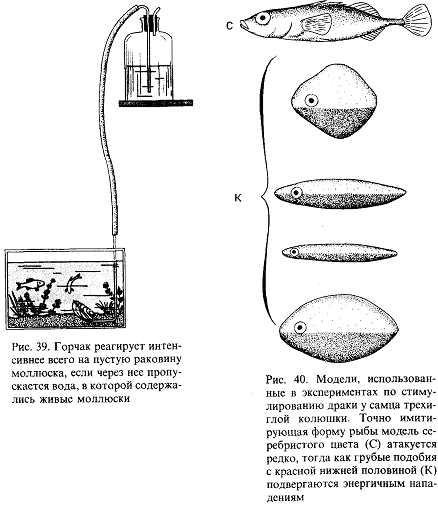
Эти стимулы анализировались у многих видов в экспериментах с макетами. Самец трехиглой колюшки хотя и проявляет некоторую враждебность к любым проплывающим мимо рыбам, особенно нетерпим к самцам своего собственного вида. Модели вызывают у него такую же реакцию, только если у них красное “брюхо”. Ярко-синие глаза и голубоватая спина почти не усиливают эффекта, а форма и размер могут варьировать в очень широких пределах. Сигарообразный макет с одним только глазом, но красной нижней половиной вызывает гораздо более интенсивную атаку, чем точно воспроизводящая форму модель или даже только что убитая колюшка без красной окраски (рис. 40). Размер значит так мало, что все наблюдавшиеся мной самцы “атаковали” даже красные почтовые фургоны, проезжавшие примерно в ста метрах от них, т. е. поднимали свои спинные шипы и неудержимо стремились догнать автомобиль, в конечном итоге, естественно, натыкаясь на стеклянную стенку. Когда фургон двигался мимо лаборатории, вдоль огромного окна которой стояли в ряд двадцать аквариумов, все самцы бросались к “оконной” стороне своего жилища и провожали фургон от одного ее угла до другого. Модели размером втрое больше колюшки вызывали сходную атаку, пока не придвигались слишком близко, но не подвергались нападению на самой территории особи. Создается впечатление, что важен угол, под которым виден объект; это объясняет, почему рыбы бросаются на удаленную красную машину.
Помимо цвета атаку может вызвать поведение. Самец колюшки, замечая издали соседа, часто принимает угрожающую позу—любопытное вертикальное положение головой вниз (рис. 41).
Рис. 41. Угрожающая поза самца трехиглой колюшки
Боком или даже брюхом он поворачивается к противнику и оттопыривает один или два брюшных шипа. Эта поза приводит других самцов в бешенство, и в эксперименте можно усилить нападения на макет, предъявив его именно в таком положении.
Сходные наблюдения проведены на зарянке. Когда самец “застолбил” территорию, появление на ней сородича вызывает нападение на него или угрозу. Лак показал, что основным пусковым фактором служит здесь красная грудь птицы. Когда он помещал на занятую территорию чучело зарянки, даже мелкого пучка красных перьев было достаточно, чтобы вызвать угрожающую позу хозяина (рис. 42). Если в случае колюшки очень грубая красная модель действовала эффективнее, чем совершенная по форме, но серебристая по

Рис. 42. Эксперимент на выявление стимулов, провоцирующих драку у зарянки. Чучело неполовозрелой зарянки с бурыми перьями на груди (слева) атакуется редко, а пучок красных перьев (справа) вызывает угрожающую демонстрацию
цвету, для зарянки эти несколько перьев значат больше, чем чучело неполовозрелого сородича, совпадающее по облику со взрослой птицей, но с бурой, а не с красной грудью. Сходство функций красного брюха самца колюшки и красной груди зарянки поразительно. Мы еще увидим, что аналогичные сигнальные системы конвергентно развились и у животных других групп.
Однако в случае зарянки сигналы не только зрительные. Птицы слышат друг друга на гораздо большем расстоянии, чем видят. В частности, песня зарянки возбуждает владельца территории и бросает его на поиски певца. Таким образом, сражение развивается в два этапа: сначала самец летит в направлении, откуда доносится пение соперника, затем оглядывается и, получив дополнительный стимул от его красной груди, принимает угрожающую позу или нападает.
У многих других видов песня выполняет ту же функцию. Это “символ мужественности”, провоцирующий владельца территории на драку. Как уже говорилось выше, идентичная символика, демонстрируемая самцом на своей собственной территории, изгоняет прочь незваного гостя. Это легко наблюдать в поле без всяких экспериментов. Часто бывает так, что поющего самца, укрывшегося в ветвях дерева или кустарника, не заметно, но реакция прочих птиц на его пение очень сильная и характерная. Особи, пролетающие сквозь чужие владения, как бы олицетворяют нечистую совесть, в то время как владельцы собственных территорий—справедливое негодование.
У серебристой чайки оперение обоих полов одинаковое, но агрессивны главным образом самцы и в отношении других самцов своего вида. Они не поют и не издают других звуков, как-то по-особому влияющих на соперников. Ярко окрашенных частей, провоцирующих нападение, у них тоже нет. Привлекают внимание других самцов и вызывают их враждебность в основном угрожающие позы и гнездостроительные движения.
Другие виды напоминают колюшку тем, что провоцирующим сигналом служит для них броская окраска соперника. Это можно, например, наблюдать у золотого дятла, самцы которого отличаются черными пятнами у углов рта (так называемыми усами), отсутствующими у самок (рис. 43). Если отловленной самке подрисовать такие усы, она будет атакована собственным партнером. Если их смыть, мир в паре восстановится.
Самец волнистого попугайчика отличается от самки цветом восковицы на клюве: у него она голубая, у той — бурая (рис. 44). Самки с перекрашенной в голубой цвет во-сковицей атакуются самцами.
Наиболее любопытная параллель обнаружена в такой непохожей на рассмотренные выше группе, как головоногие моллюски. Самцам обыкновенной каракатицы в период спаривания свойственна удивительная зрительная демонстрация. Встречая другую каракатицу, они показывают ей широкую сторону своих рук, на которых за счет координированного действия хроматофоров возникает яркий узор из темно-пурпурных и белых полос (рис. 45). Реакцией на это будет драка самцов; эксперименты с гипсовыми моделями показали, что демонстрация действует зрительно; для нападения важны и цвет, и форма узора.
Ящерицы в этом смысле очень близки к каракатицам. Особые движения самца служат для демонстрации его специфической окраски. У американской заборной игуаны окраска спины покровительственная, однако брюхо самцов голубое. Этот цвет незаметен, пока рептилия его не демонстрирует, встречаясь весной с соперником. Самец поворачивается к нему боком и сжимает тело с боков, так что голубое брюхо становится хорошо заметным (рис. 46). Изменяя окраску заборных игуан и самок с помощью лаков, Нобл показал, что агрессивность у охраняющих территорию самцов провоцирует именно голубой цвет брюха.
До сих пор рассматривались примеры стимулов, определяющих главным образом время драк. Как правило, они одновременно указывают цель нападения. Однако, как и в случае брачного поведения, следует различать синхронизирующую и ориентирующую функции, поскольку некоторые стимулы связаны только с одной из них. Например, самки уток особыми движениями и криками побуждают своих партнеров нападать на других самцов. При этом крики просто стимулируют агрессивность, тогда как движениями головы самка указывает на объект атаки. Это легко наблюдать в случае ручных и полуприрученных крякв, обитающих в парках: самка плывет от “пристающего” самца к собственному партнеру, неоднократно оборачиваясь через плечо в сторону чужака.
Приведенные выше примеры касаются и третьей проблемы — репродуктивной изоляции, т. е. ограничения драк представителями одного и того же вида. Опять же, как и в случае сигналов, управляющих брачным поведением, стимулы, провоцирующие драку, видоспецифичны и весьма своеобразны даже у близкородственных видов, если они живут в одном и том же местообитании. Однако создается впечатление, что межвидовые драки не были так строго исключены в процессе эволюции, как межвидовое скрещивание. Редкие стычки наблюдаются между особями разных, но внешне сходных видов. “Ошибочное” нападение объясняется присутствием у чужого вида каких-либо знаковых стимулов, обычно вызывающих атаку. Однако в некоторых случаях драка явно затевается с другим видом из-за конкуренции за один и тот же “необходимый объект”. Так, известно, что скворцы и древесные овсянки отгоняют другие виды птиц от своих гнездовых дупел.
Живущие группами животные часто дерутся не только за самку или территорию. Они могут вступать в столкновения из-за корма, удобного места и по другим причинам. В таких случаях число стычек часто снижается в результате научения. Каждый индивид на собственном приятном или горьком опыте узнает, которого из сородичей следует избегать, поскольку тот сильнее, а кто достаточно слаб, чтобы его можно было испугать. В результате возникает “порядок клевания”, при котором каждый член группы знает свое место. Один индивид становится тираном, подавляющим всех прочих. Кто-то подчиняется только ему. “Третий номер” подавляет всех, кроме двух вышестоящих, и т. д. Такой иерархический порядок известен у различных птиц, млекопитающих и рыб. Его легко наблюдать в курятнике.
Иерархия—одно из средств сокращения количества настоящих драк. Особи, не научившиеся быстро избегать старших по рангу, оказываются в неблагоприятном положении как из-за того, что их больше бьют, так и из-за повышенной уязвимости для хищников во время стычек.
Поведение, ведущее к установлению иерархического порядка, имеет несколько интересных аспектов. Лоренц наблюдал у галок, что, если самку низшего ранга выбирает в партнерши самец, стоящий выше по иерархической лестнице, эта самка немедленно повышается до него в ранге, т. е. все особи, стоящие ниже самца, начинают ее избегать, хотя некоторые из них раньше занимали высшее положение.
В американской литературе можно найти множество ценной информации по проблемам иерархии. Однако многими авторами она рассматривается как единственный принцип социальной организации. Это не совсем так; иерархические отношения—лишь один из множества типов социальных взаимодействий.