
- Орангутаны научились использовать орудия с выгодой для себя [2019-03-22]
- Мозг собак отличил настоящие слова от тарабарщины [2019-03-22]
- Какаду Гоффина выклевали палочки из картонки и использовали их в быту [2019-03-22]
- Орангутанов уличили в создании крючков [2019-03-22]
- Любовь самок гуппи к ярким самцам объяснили генами и освещением [2019-03-22]
- Орангутаны рассказывают друг другу о прошлом [2019-03-22]
- Блеск и нищета этологии [2018-05-03]
- Почему хозяева похожи на своих собак? [2018-03-15]
- Интервью с Анатолием Протопоповым [2019-02-13]
- Цирки: развлечение vs мучение [2018-12-15]

David M. Buss University of Texas at Austin Department of Psychology
Abstract
Modern humans have inherited the mating strategies that led to the success of their ancestors. These strategies include long-term mating, short-term mating, extra-pair mating, mate poaching, and mate guarding. This article presents empirical evidence supporting evolution-based hypotheses about the complexities of these mating strategies. Since men and women historically confronted different adaptive problems in the mating domain, the sexes differ profoundly in evolved strategic solutions. These differences include possessing different mate preferences, different desires for short-term mating, and differences in the triggers that evoke sexual jealousy. The study of human mating is one of the “success stories” of evolutionary psychology.
Strategies of Human Mating
No adaptive domain is more central to reproduction than mating. Those in our evolutionary past who failed to mate failed to become ancestors. Modern humans are all descendants of a long and unbroken line of ancestors who succeeded in the complex tasks involved required to mate successfully. As their descendants, modern humans have inherited the adaptations that led to the success of their ancestors.
Successful mating requires solutions of a number of difficult adaptive problems. These including selecting a fertile mate, out-competing same-sex rivals in attracting a mate, fending off mate poachers (those who try to lure one’s mate away), preventing the mate from leaving, and engaging in all of the necessarily sexual and social behaviors required for successful conception to take place. As a consequence of the number and complexity of mating problems humans have recurrently faced over the long expanse of human evolutionary history, it is
David M. Buss, Department of Psychology, University of Texas, Austin, Texas 78712. E-mail: dbuss@psy.utexas.edu
reasonable to anticipate that humans have evolved a large and complex array of adaptations specifically dedicated to the task of mating.
Nowhere do people have an equal desire to mate with all people. Everywhere, some people are preferred as mates, others shunned. Desires are central to all facets of mating. They determine who we are attracted to, and who is attracted to us. They influence which attraction tactics will be successful (those that fulfill desires) and which attraction tactics will fail (those that violate desires). Successful mate retention tactics involve continuing to provide resources that fulfill the desires of a mate. Failure to fulfill these desires causes breakup and divorce. At every step of the mating process, from mate selection to mate expulsion, desires determine the ground rules.
Sexual Selection and Parental Investment
Although Charles Darwin (1859) recognized that survival was central to the evolutionary process, many natural phenomena he observed seemed to baffling on the theory of “survival selection.” He noticed phenomena such as the brilliant plumage of peacocks, the flamboyant feathers of cardinals, and the enormous antlers of deer. How could these metabolically costly features possibly have evolved? Many seemed like open lures to predators, and hence detrimental to survival. Darwin also noticed that males and females of many species appeared to be different in size and shape. Male elephant seals, for example, weight roughly 4,000 pounds; female elephant seals weigh only 1,000 pounds. Among baboons, males are twice the size of females. Among humans, males are 12 percent taller than females, on average. Since both sexes have faced roughly the same survival problems, why would they differ in size and morphology? And what could account for variation on the degree of sexual dimorphism across species?
Darwin’s answer to these empirical puzzles was the theory of sexual selection (Darwin, 1858, 1871). The theory of sexual selection dealt with the evolution of characteristics due to mating, rather than survival, advantage. Darwin described two component processes through which sexual selection could take place. In the first, called intrasexual competition, members of one sex (often, but not always, the males) engaged in competitive battles with each other. Two stags locking horns in combat is an excellent example of intrasexual competition. The victors in these battles gain preferential sexual access to females. The losers fail to mate. The qualities that lead to success in same-sex contests, therefore, are passed down in greater numbers (assuming that these qualities are partly heritable). Whatever qualities are linked with losing either fail to get passed down. Evolution, that is change over time, occurs as a result of the differential reproduction of the winners and losers in same-sex contests.
It is important to note that intrasexual competition need not always direct physical combat. Males in some species compete for position in the status or dominance hierarchy through non-physical means, and position in the hierarchy can be linked with preferential access to mates (e.g., Betzig, 1986; Buss, 1994/2003). Males in other species scramble for access to territory, and access to territory can be linked to preferential access to mates. The key point is that whatever qualities lead to success in intrasexual competition are passed on in greater numbers, whether the competition is physical combat, maneuvering for position in the hierarchy, or scramble for access to certain resources. The result is evolution through sexual selection.
The second process through which sexual selection occurs is intersexual selection. This process involves the preferences of members of one sex for members of the opposite sex who possess certain qualities. Hypothetically, if all women preferred to mate with men who had red hair, those with red hair would have a mating advantage. Over time, we would witness an increase in the frequency of red-headedness in the population. The key point is that the desires of one sex for certain qualities in a mate can create evolutionary change - either an increase in the frequency of desired qualities or a decrease in the frequency of undesired qualities. Although Darwin called this process “female choice,” it is clear that in many species, and certainly in humans, males also exert considerable mate choice. Theoretically, the mate preferences of one sex can determine over evolutionary time the domains in which the opposite sex competes (see Figure 1). If females desire males who build sturdy nests, then males will compete with each other to construct nests that embody the female preferences. Conversely, the domains in which one sex competes can influence the evolution of mate preferences in the other sex. If males compete with other males to monopolize resource-rich territories, for example, females might subsequently evolve a mate preference for males with large territories. Thus, mate preferences and patterns of intrasexual competition can co-evolve, each influencing the other.
Figure 1. Sexual Selection Subsumes the Processes of Intrasexual Competition and Intersexual Selection
Sexual Selection
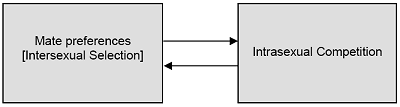
The mate preferences of one sex determine the domains in which the opposite sex will compete. The ways in which one sex competes can influence the evolution of mate preferences.
Darwin’s theory of sexual selection was initially designed to explain the various empirical puzzles he had observed—things like the brilliant plumage of peacocks (preferred by peahens) and the larger size of males in some species (explained by the advantage that size gives males in intrasexual competition, or by female preferences for males who are large). But many puzzles remained. Darwin observed that females were often the choosy sex, but he did not know why. He also observed that males were often the competitive sex, but he did not know why. Roughly a century would pass before evolutionary biologists devised a powerful theory to explain what determines which sex will compete and which sex will exercise choice, that is, what drives the operation of the two component processes of sexual selection.
Trivers’s (1972) answer to these questions was the theory of parental investment. According to this theory, the sex that invests more in offspring would be more choosy about mates. In species with internal female fertilization, the greater parental investment by females makes them a valuable reproductive resource. Gestating, bearing, and breast feeding a child, for example, are costly endeavors. Elementary economics tells us that those who hold valuable resources do not give them away indiscriminately. Evolution favored women who were highly selective about their mates. Women who were not choosy would have suffered lower reproductive success. Those who engaged in careful mate selection, for example preferring a man who would stay around, invest in her, and protect her children, enjoyed reproductive benefits. The more an individual devotes effort to parental investment, according to Trivers, the greater the benefits of exercising careful mate choice. The sex that invests less in offspring, according to this theory, should be more competitive with each other for access to the high-investing sex. In summary, the relative investment of the two sexes drives the operative components of sexual selection, with the high investing sex being selected to be the most choosy and the lower investing sex being the most competitive with members of their own sex for mating access to the more valuable high-investing sex.
The Menu of Human Mating Strategies
One of the intriguing features of human mating is that it cannot be characterized by a single strategy. One strategy on the menu is long-term committed mating, often, but not always, characterized by a formal public commitment such as marriage. In long-term mating, both sexes typically invest heavily in any resultant offspring. As a consequence, and in accordance with the theory of parental investment, sexual selection has likely fashioned in both sexes high levels of choosiness or selectivity. Poor long-term mate choices would have been costly for either sex because they would have risked wasting their heavy investments.
Not all mating, however, lasts a long time. Human matings can last a few years, a few months, a few weeks, a few days, or even a few minutes. One end of this temporal continuum may be called short-term mating (Buss & Schmitt, 1993). This temporal dimension turns out to be critical to many components of mating, perhaps none more central than the qualities desired. Furthermore, humans display remarkable creativity in their ability to mix and match mating strategies. It is not uncommon, for example, for a person to engage in one long-term committed mateship with heavy investment in children, while simultaneously pursuing an extramarital affair, or series of affairs, on the side.
Humans, in short, are neither solely monogamous, nor solely promiscuous; neither polygynous nor polyandrous. Which strategies from the menu a particular person chooses is heavily dependent on circumstances. These include the sex ratio in the mating pool (i.e., the ratio of males to females), a person’s mate value, and even prevailing cultural norms (Buss, 2004). These issues are discussed later, but first, we must outline the central desires of men and women in their pursuit of long-term and short-term mates.
Qualities Desired in a Marriage Partner
Because women have a large obligatory parental investment to produce children, and therefore are predicted to be discriminating in their mate choice, the next key question is: discriminating about which qualities? Potential mates vary in thousands of ways, from physical prowess to speed of hair growth. Adaptationist thinking provides a guide to hypotheses about the evolution of what women want, namely those characteristics that reliably led to an increase women’s reproductive success. These include selecting a mate who (1) is able to invest resources in her and her children, (2) is willing to invest resources in her and her children, (3) is able to physically protect her and her children, (4) is willing to physically protect her and her children, (5) show promise as a good parent, and (6) will be sufficiently compatible in goals and values to enable strategic alignment without inflicting too many costs on her and her children (Buss, 1994/2003).
In a large-scale cross-cultural study, Buss and his colleagues (Buss, 1989; Buss et al., 1990) explored how much women and men desired each of 32 qualities in a potential long-term mate. The study involved samples from 37 cultures located on six continents and five islands. The samples included Gujarati Indians, Estonians, mainland Chinese, Santa Catarina Brazilians, and South African Zulu. The sample from Croatia was obtained from Zagreb. The total sample size was 10,047, with an average of 272 from each of the 37 cultures.
Cultures varied tremendously in the value placed on some characteristics. The desire for chastity or virginity (lack of prior sexual intercourse) proved to be the most cross-culturally variable, as shown in Figure 2. Mainland Chinese placed tremendous value on virginity, as did participants living in India and Iran. At the
other end of the scale, the Dutch placed little importance on chastity. Those from Western Europe, especially Denmark, Sweden, Finland, and German also placed little importance on virginity. Overall, 62% of the cultures showed a significant sex difference, always in the direction of men valuing virginity more than women. There were no reversals of this pattern. On the other hand, 38% of the cultures showed no significant sex difference. These findings suggest that the importance placed on chastity is highly susceptible to cultural input, with cultures differing from each other both in the absolute value placed on chastity as well as in the presence of absence of sex differences.
Figure 2. The Rating Scale Ranged from 3 (Indispensable) to 0 (Irrelevant or Unimportant) in a Marriage Partner. Sex Differences are Statistically Significant (p < .05) for india, japan, usa, and croatia, but not for china or nl (the netherlands)
Chastity: No previous experience
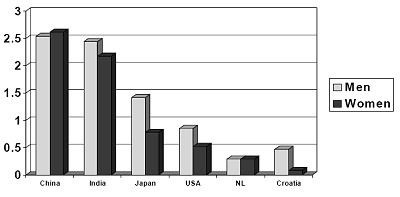
China India Japan USA NL Croatia
Many characteristics were universally desired by both sexes. Worldwide, women and men wanted mates who were intelligent, kind, understanding, dependable, and healthy. Similarly, mutual attraction/love emerged as one of the most valued qualities in a spouse worldwide. Both sexes also valued potential spouses who were similar to themselves in their political orientation and religious beliefs.
Despite these cultural variations and universal commonalities, women and men differed across the globe on their desire for some qualities, precisely as predicted in advance by the evolutionary hypotheses. Women, significantly more than men, desired “good financial prospect” (see Figure 3). Women also tended to value qualities that are known to be linked to resource acquisition, such as ambition, industriousness, social status, and somewhat older age.
Figure 3. Rating Scale Ranged from 3 (Indispensable) to 0 (Irrelevant or Unimportant) in a Marriage Partner. Sex Differences are Statistically Significant (p < .05) in all cultures
Good financial prospect
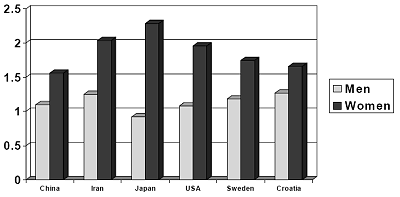
China Iran Japan USA Sweden Croatia
Men, significantly more than women, desired partners who are “good looking” and “physically attractive.” Physical appearance, as voluminous research has shown, provides a wealth of cues to a woman’s health, fertility, and reproductive value (see Figure 4). Contrary to long-held beliefs among social scientists, standards of beauty are not arbitrary or infinitely culturally variable. Evolutionary psychology provides a powerful theory for the evolution of standards of beauty-whatever observable cues are linked with fertility (immediate probability of conception) or reproductive value (future reproductive potential) will evolve to become part what humans find attractive in female. These include cues to youth, such as full lips, smooth skin, lustrous hair, and a low ratio of hips to waist (WHR). They also include cues to health, such as clear skin, absence of sores, white teeth, and symmetrical features. Beauty, in short, is in the adaptations of the beholder, and men value physical appearance because of the wealth of information it provides about a woman’s youth, health, and hence reproductive capacity.
Figure 4. Graph Depicts the Logic of the Hypothesis of Evolved Standards of Beauty
Logic of the Evolution of Standards of Attractiveness
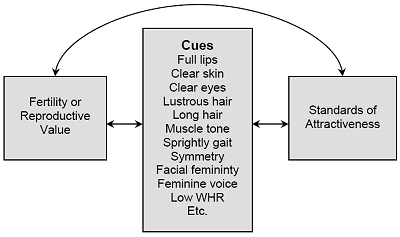
Observable cues such as full lips and clear skin are hypothesized to become part of evolved standards of female attractiveness because of their link with fertility (immediate probability of reproduction) or reproductive value (future reproductive potential)
Men universally wanted mates who were younger than themselves, confirming the hypothesis that men desire fertility cues (see Figure 5). Evolutionary models have predicted that what men desire is not youth per se, but rather features of women that are associated with reproductive value or fertility. This perspective leads to a counterintuitive prediction when it comes to the age preferences of adolescent males: teenage males are predicted to prefer women who are slightly older then they are, contrary to the typically observed pattern of men desiring younger women. This prediction is based on the fact that women slightly older than these teenaged boys have slightly higher fertility than women their own age or women who are younger (Kenrick, Keefe, Gabrielidis & Cornelius, 1996).
The findings of the Kenrick et al. (1996) studies confirmed this counterintuitive prediction. Although teenage males were willing to accept dates with women who were slightly younger, they found women a few years older to be the “most attractive.” Interestingly, this finding occurs despite the fact that these older women express no interest at all in dating younger men. Taken together, the cumulative findings suggest that men’s age preferences exist, at least in large measure, because of the historically recurring link between a woman’s age and her fertility.
Figure 5. Participants Reported in Years Their Preferred Age Difference Between Self
and Spouse, Expressed in Number of Years Older or Younger
Age difference preferred between self and spouse
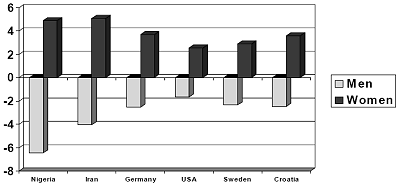
Nigeria Iran Germany USA Sweden Croatia
Men universally preferred younger partners, whereas women universally preferred older partners. In Croatia, for example, women wanted men nearly four years older then themselves, whereas men wanted women roughly two and half years younger than themselves. The sex differences are statistically significant (p < .001) in all cultures.
In summary, universal sex differences occurred in precisely those domains predicted to involve sex-linked adaptive problems, notably the selection of mates who have the ability to invest resources (women prefer more than men) and mates who display cues to youth and beauty, known signals of fertility (men prefer more than women). Despite these universal sex differences, most mate preferences show great similarity between the sexes (e.g., kind and understanding, intelligent, healthy), and there are also important cultural differences in the desires (e.g., chastity).
It is important, of course, to obtain independent confirmation of these findings from alternative methods that do not rely on expressed preferences. And indeed, many alternative methods support the validity of the methods used to obtain expressed preferences. A study of actual marriages in 29 different cultures, for example, confirmed that men do choose younger women (Buss, 1989). Grooms were older than brides in each one of the 29 cultures, with an average age difference of three years. Furthermore, as men get older, if they get divorced and remarry, they tend to marry women who are increasingly younger than they are. The age gap is three years at first marriage, five years at second marriage, and eight years at third marriage (Buss, 1994/2003).
Studies of the response rates to personals ads also confirm the results found with expressed preferences. Women mentioning physical attractiveness and young age as part of their self-description in their ads receive significantly higher response rates than women who are older or who fail to mention anything about their physical attractiveness. Conversely, men who mention excellent financial resources in their self-descriptions in their ads received a higher response rate from women than men who fail to mention this attribute (Baize & Schroeder, 1995).
Finally, studies of the behavioral tactics that men and women use to attract mates, retain mates, and derogate their rivals all correspond closely to the expressed desires of the opposite sex. Women, for example, tend to put more effort into appearance enhancement in mate attraction and mate retention, and when they derogate their rivals they focus on the rival’s physical flaws (e.g., mentioning that the other woman’s thighs are heavy). Conversely, men tend to display and bestow resources on the women they are trying to attract and retain. They tend to denigrate their rivals by impugning the rival’s professional prospects, such as mentioning that he rival is lazy, lack ambition, or lacks clear goals in life (see Buss, 2004, for detailed summaries of these studies). When men and women attempt to deceive each other, they do so precisely along the lines of the desires expressed by the opposite sex (Tooke & Camire, 1991).
It is worth noting that, conceptually, we do not expect a perfect correspondence between expressed desires and actual mating behavior for the simple reason that people cannot always get what they want. A person’s own level of desirability, for example, will limit the ability to translate ideal mate preferences into an actual mating. Most people must settle for someone who is less than what they ideally want. Nonetheless, the available evidence converges from a variety of different methods that these fundamental desires differ for men and women and affect actual mating behavior in precisely the ways predicted.
Desires in Short-Term Mating
Trivers’s (1972) theory of parental investment provides a powerful basis for predicting sex differences in the pursuit of short-term matings. Men, more than women, are predicted to have evolved a greater desire for casual sex. The same act of sex that causes a woman to invest nine months in pregnancy obligates the man to little or no investment. Over a one-year period, an ancestral man who managed to have short-term sex with dozens of women would have caused many pregnancies. An ancestral woman who had sex with dozens of men in the same year would produce only a single child. The reproductive benefits to men of short-term mating, in sum, would have been a direct increase in the production of children. A married man with two children, for example, would have increased his reproductive success by 50 percent by one short-term copulation or affair that resulted in conception and birth.
The empirical evidence for a sex difference in desire for short-term mating is extensive, supported by hundreds of scientific studies. When asked how many sex partners they would ideally like, men state that they would like 18 in their lifetime, whereas women average around 4.5, as shown in Figure 6 (Buss & Schmitt, 1993).
These large sex differences have been replicated using different statistical methods of calculating central tendency (e.g., medians rather than means) on samples diverse in age (Schmitt et al., 2003).
Figure 6. Number of Sexual Partners Desires
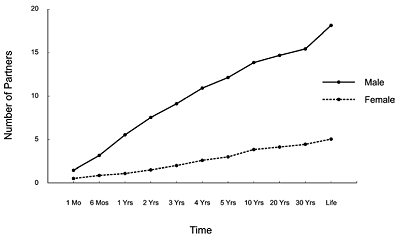
Participants recorded in blank spaces how many sexual partners they ideally would like to have for each specified time interval (data from Buss & Schmitt, 1993).
Another psychological solution to the problem of gaining sexual access to a variety of partners is to let little time elapse between meeting a desired female and seeking sexual intercourse. The less time a man permits to elapse before seeking sexual intercourse, the larger the number of women he can succeed in copulating. In one study that has been extensively replicated, men and women rated how likely they would be to consent to sex with someone they viewed as desirable if they had know the person for only an hour, a day, a week, a month, and so on. Both men and women say that they would probably have sex after knowing a desirable potential mate for five years (see Figure 7). At every shorter interval, men exceeded women in the reported likelihood of having sex.
Figure 7. Participants Indicated How Likely They Would Be to Have Sex With Someone They Found Attractive as a Function of How Much Time They Had Known the Individual
Likelihood of agreeing to have sex with someone you find attractive as a function of time known
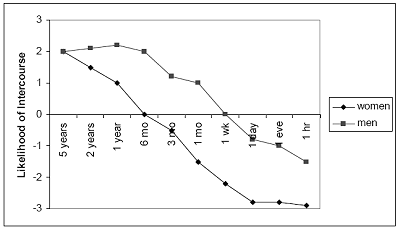
The scale ranged from +3 (highly likely) to -3 (highly unlikely).
A behavioral study confirmed this large sex difference (Clarke & Hatfield, 1989). Men and women experimenters approached total strangers on a college campus, and said “Hi, I’ve been noticing you around campus, and I find you very attractive.” Then they asked one of three questions: Would you go out on a date with me? Would you go back to my apartment with me? Would you have sex with me? The experimenters recorded the percentage who agreed to each request, and also any verbal comments they made.
Of the women approached by the male experimenters, 50% agreed to go out on a date with him; 6% agreed to go back to his apartment; and 0% agreed to have sex. Some women who were asked for sex were insulted, and some thought is bizarre. Of the men approached by the female experimenters, 50% agreed to go out on a date, similar to the women’s responses. However, 69% agreed to go back to her apartment. And 75% agreed to have sex with her. Of the men who declined the sexual request, some were apologetic, citing a prior commitment with parents of a fiancé. These sex differences have been replicated in subsequent studies (see Buss, 2000, for a summary).
In summary, the evidence is strong that men have evolved psychological mechanisms dedicated to solving the complex problems posed by success at short-term mating. These include a desire for sexual variety, the tendency to let little time elapse before seeking sexual intercourse, and the behavioral willingness to consent to sex with strangers. In addition, men appear to lower their standards dramatically in the context of short-term mating (Buss & Schmitt, 1993); show a marked decrease in attraction to a sex partner immediately following sexual intercourse, perhaps to facilitate a hasty post-copulation departure (Haselton & Buss, 2001); report exaggerating the depth of their feelings to gain sexual access (Haselton, Buss, Oubaid & Angleitner, 2005); and report that they would have an extramarital affair if they knew that no one would find out (for reviews of this evidence, see Buss, 1994/2003).
Women’s Short-Term Mating Strategies
Although the empirical evidence is clear that men, far more than women, have a great desire for a variety of sex partners, men could never have evolved such a desire in the absence of willing women (barring deception or forced intercourse). Indeed, mathematically, the mean number of sex partners for men and women must be identical, assuming an equal sex ratio in the population. Ever time a man has sex with a woman with whom he has not previously had sex, a woman is simultaneously having sex with a man with whom she has never had sex.
Perhaps because the evolutionary logic for men having evolved a strong desire for sexual variety is so clear - namely, an increase in direct reproductive output the evolutionary logic for women having evolved a short-term mating psychology has been relatively neglected. The puzzle is deepened by the fact that short-term mating often carries substantial costs for women. Women, more than men, risk damage to their reputations, a lowering of perceptions of their mate value, and if mated, the possibility of violence at the hands of a jealous boyfriend or husband. Given these costs, it is unlikely that selection would have forged a female short-term mating psychology in the absence of substantial benefits that outweigh those costs.
In an effort to explore what those benefits might be, Greiling and Buss (2000) extracted from the literature and formulated a number of hypotheses about potential benefits that women could obtain from short-term mating. These include resource hypotheses (e.g., immediate resource accrual), genetic hypotheses (e.g., producing more genetically diverse offspring), mate switching hypotheses (e.g., using a short-term mating as a means to exit a poor mateship), mate skill acquisition hypotheses (e.g., clarifying mate preferences), and mate manipulation hypotheses (e.g., deterring a partner’s future infidelity).
Greiling and Buss (2000) conducted a series of four empirical studies to identify which hypotheses appeared promising and which did not. Although limited in scope, these studies were designed to examine (1) the perceived likelihood that a woman would receive particular benefits through a short-term mating; (2) the perceived magnitude of benefits if received; (3) the contexts in which women engage in short-term mating; and (4) individual differences among women in proclivity to engage in pursue short-term matings in their perceptions of benefits. Below are reported only the result of short-term extra-pair mating (EPC).
The hypotheses that received the most empirical support across studies were those of resource acquisition and mate switching. For example, women judge it to be highly likely that they will receive jewelry, money, free dinners, or clothing by engaging in short-term mating. Furthermore, a critical context if which women consider short-term affairs is when the partner cannot hold down a job. Women who actively engage in short-term mating, in contrast to their more monogamous counterparts, judge the resource benefits to be “more beneficial.”
The hypothesized “mate switching function” of women’s short-term mating would, of course, only apply to context in which the short-term mating is an affair or an extra-pair copulation (EPC). Contexts in which women judge it to be highly likely that they will have an affair include “feeling that she could find someone with whom she is more compatible than her current partner.” Furthermore, women perceive it to be highly beneficial to discover a sexual partner who is interested in making a commitment to them, willing to spend a lot of time with them, and able to replace her current partner.
Other research, however, also supports the “good genes” hypothesis (Gangestad & Thornhill, 1997). Symmetry of features and masculine facial features, for example, are hypothesized to be markers of good genes. Women show a special preference for men who are symmetrical and masculine when they are ovulating - precisely the time when they are most likely to conceive a child (Gangestad et al., 2005). In summary, mate switching, resource acquisition, and good genes are three strong contenders for the evolved functions of women’s short-term mating. Further research, of course, is needed to test these hypotheses.
The existence of already mated women who sometimes engage in sexual intercourse with other men (extra-pair mating) points to an adaptive problem that men face-the presence of mate poachers.
The Strategy of Mate Poaching
Mate poaching may be defined as behavior designed to lure someone who is already in a romantic relationship, either temporarily for a brief sexual liaison or more permanently for a long-term mating. Until recently, practically nothing was known scientifically about the phenomenon of mate poaching. According to one study (Schmitt & Buss, 2001), mate poaching turns out to be a prevalent phenomenon. Using a relatively mature sample of American participants, averaging 41 years of age (range = 30 – 65), 60% of the men and 53% of the women reported having attempted to poach someone as a long-term mate who was already in an existing committed relationship. The comparable figures for attempting to attract an already-mated person for a short-term sexual liaison were 60% for men and 38% for women.
The majority of this sample also reported being recipients of mate poaching attempts by others while they were in a committed romantic relationship. These figures for the long-term mating context were 93% for men and 82% for women. Eighty-seven percent of the men and 94% of the women reported being recipients of mate poaching attempts for brief sexual matings.
Attempted mate poaching is one issue; successful mate poaching is another. When asked whether they have been successfully lured away from an existing relationship, 67% of the men and 41% of the women responded affirmatively for the long-term context. And 40% of the men and 31% of the women report having been successfully seduced by a mate poacher for a short-term sexual liaison. Similar findings have been obtained cross-culturally in samples from Israel, Turkey, Greece, Croatia, Slovenia, Poland, Portugal, Germany, France, England, and Canada (Schmitt et al., 2004).
It is likely that mate poaching is an evolved mating strategy for the simple reason that desirable mates attract many suitors, and typically end up in mating relationships. Thus, in order to obtain a desirable mate, it is often necessary to seek those who are already “taken.” The unique mate poaching tactics, such befriending the couple and waiting in the wings for an opportunity, or attempting to drive a wedge in the relationship, reveals the lengths to which people will go in order to mate poach.
Strategies of Mate Guarding
Infidelity and mate poaching as mating strategies pose serious adaptive problems for the “victims,” that is the mates of the individuals who are committing infidelity or being tempted by a mate poacher. If these mating strategies have recurred over the long course of human evolutionary history, as the evidence suggests, the principle of co-evolution dictates that strategies will almost certainly evolve to defend against these problems and the costs they impose. One possible solution involved the evolution of jealousy (Buss et al., 1992; Daly, Wilson & Weghorst, 1982; Symons, 1979). Jealousy is an emotion that is activated whenever there is a threat to a valued relationship (Daly et al., 1982). Threats can come in many forms, such as the loss of a partner’s sexual, financial, or emotional resources to a rival. Threats can come from within the relationship from a partner who might have the urge to stray, or from outside the relationship in the form of mate poachers attempting to lure a partner away.
Over the past decade, a substantial amount of research has been devoted to exploring jealousy as an evolved solution to the problems of infidelity and mate poaching (see Buss, 2000, for an in-depth discussion). The specifics of the adaptive problems differ for men and women, according to evolutionary theorists (Daly et al., 1982). Because in humans fertilization occurs internally within women, men can suffer a lack of certainty in their paternity. In contrast, women are always 100% certain that their offspring are their own. Sexual infidelity, of course, is the event that can compromise a man’s paternity in offspring. Although women have never confronted the problem of maternity uncertainty, an infidelity by a woman’s mate can be extremely damaging. The woman whose husband is unfaithful risks losing his time, resources, and commitments, all of which could get channeled to a rival female and her children. For these reasons, evolutionary theorists have predicted that men, more than women, would get upset about signals of sexual infidelity. In contrast, women, more than men were predicted to get upset about signals of emotional infidelity, since emotional involvement is a leading indicator of the diversion of these resources and commitments (Buss et al., 1992).
Dozens of empirical studies, using a variety of methodologies, have now been conducted to test for this sex difference (Buss & Haselton, 2005). In one study, participants were asked to imagine that their romantic partner had become both sexually and emotionally involved with someone else (Buss et al., 1999). Then they were asked to indicate which aspect of the betrayal was more upsetting. In an American sample, 61% of the men, but only 13% of the women judged the sexual infidelity aspect of the betrayal to be the most upsetting. Conversely, only 39% of the men, but 87% of the women, judged the emotional attachment to the other person as more upsetting. Similar sex differences have been obtained in Korea and Japan (Buss et al., 1999), China (Geary et al., 1995), and Sweden (Wiederman & Kendall, 1999). In studies of memory, men can more easily recall cues to sexual infidelity, whereas women can more easily recall cues to emotional infidelity.
In summary, men and women differ, as originally predicted in advance by evolutionary theorists, in the weighting given to the events that activate jealousy. Men, more than women, tend to become extremely distressed over signals of sexual infidelity; women more than men tend to become more distressed over signals of emotional infidelity. Of course, both sexes typically get extremely upset by both forms of infidelity, as they should given that both forms threaten key reproductively relevant resources. Furthermore, the two forms of infidelity are positively correlated in everyday life - people tend to become sexually involved with those with whom they are emotionally involved and vice-versa. Nonetheless, the findings are clear in supporting the original predictions about the psychological design of jealousy as an evolved defense against infidelity and the threats posed by mate poachers.
Men and women also appear to be threatened by somewhat different qualities in intrasexual rivals. Specific evolution-based predictions were testing in a cross-cultural study that included Korea, the Netherlands, and the United States (Buss et al., 2000). Korean, Dutch, and American men, more than corresponding women, reported greater distress when a rival who was interested in their partner surpassed them on financial prospects, job prospects, and physical strength. In contrast, Korean, Dutch, and American women report greater distress when a rival surpasses them on facial and bodily attractiveness. Although both sexes are equally jealous overall, the sexes differ in the weighting given to sexual versus emotional infidelity as well as in the qualities of rivals that they find threatening.
If jealousy is an evolved emotion, and the empirical evidence so far appears to support this proposition, then the next step is to explore the behavioral output of this emotion. Three different studies have explored “mate retention tactics” of men and women, using both married couples and dating couples as participants (Buss, 1988; Buss & Shackelford, 1997; Shackelford et al., 2005). Mate retention tactics are specific behaviors designed to ward off rivals or to deter a mate from straying. The specific tactics range from vigilance (e.g., He called her at unexpected times to see who she was with) to violence (e.g., He hit the guy who made a pass at her).
Married men tend to engage in especially vigorous mate retention efforts when their spouse is young in age and physically attractive. In contrast, women tend to engage in especially vigorous mate retention efforts when married to men who have good jobs, high incomes, and devote a lot of time to status striving. In addition, men and women differ in the types of mate retention tactics they use. Men, more than women, tend to display resources to their mate, as well as threaten and commit violence on intrasexual rivals. Women, more than men, tend to enhance their physical appearance as a mate retention strategy, as well as intentionally evoking their partner’s jealousy. Intentionally evoking jealousy, for example by flirting with other men and eliciting their interest, appears to be a strategy women use to increase their mate’s perceptions of their desirability (Buss, 2000).
Conclusions
Humans have evolved a complex menu of mating strategies. These include long-term committed mating, brief sexual encounters, infidelity, mate poaching, and mate guarding. Long-term mate preferences are complex, reflecting desires for many different qualities such as kindness, intelligence, mutual attraction, love, dependability, and good health. Two universal clusters of sex differences are the desire for youth and beauty (men value more than women) and the desire for a mate who has good financial prospects and elevated social status (women value more than men). These profound sex differences have been documented in studies of expressed preferences, as well as in studies of actual marriages, responses to personals ads, and tactics of mate attraction, mate retention, competitor derogation, and intersexual deception.
The empirical evidence supports the evolution-based hypothesis that men have evolved a more powerful desire for a variety of sex partners than have women. The evolutionary logic for this sex difference is straightforward - men who succeeded in securing sexual access to a variety of women would have achieved greater reproductive success than men who did not. Women, in contrast, generally cannot increase their reproductive output by having sex with many man. Nonetheless, there is a hidden side to female sexuality, and some women some of the time also pursue short-term matings. Because women’s short-term mating can be risky, it is reasonable to hypothesize that ancestral women received benefits from short-term mating that outweighed the costs. The leading hypotheses for why women engage in short-term mating, especially extra-pair mating, are the possibility of acquiring good genes that can be passed to her offspring, increasing her access to material resources, and the possibility of switching to a better mate. Nonetheless, women who cuckold their husbands historically have inflicted large reproductive cost on their regular mates. Cuckolded men risk diverting years or decades of parental resources to a rival’s offspring. The principle of co-evolution predicts that men have evolved adaptations designed to defend against the diversion of their mate’s sexual and reproductive resources.
Jealousy as an emotion has been proposed as one such evolved defense mechanism. The empirical evidence strongly supports several evolution-based hypotheses about the psychological design of jealousy. Male jealousy, more than women’s, is triggered by signals of sexual infidelity and rivals to exceed them on the qualities that women are known to want in a mate such as good financial prospects. Women’s jealousy, more than men’s, is activated by signals of emotional infidelity (and hence potential long-term diversion of commitments) as well as by rivals who exceed them on facial and bodily attractiveness.
Much more research needs to be conducted on the complexities of human mating strategies. At this point in the evolutionary psychology, however, scientists now have some of the broad outlines of the fundamentals of human mating strategies and the ways in which they are designed differently in men and women.
Additional research is needed on the context-sensitive nature of human mating strategies. Precisely which circumstances might cause a person to shift from a long-term mating strategy to a short-term mating strategy or vice-versa? Which circumstances might trigger an extramarital affair, or conversely, cause someone to forgo an alluring sexual opportunity? How do the various desires combine, given social contexts and a person’s own level of desirability, to form actual mate choices? These and other complexities of human mating are currently being explored by scientists who have grasped the centrality and importance of human mating to so many different dimensions of social living. The area of human mating is one of the true “success stories” of evolutionary psychology.
REFERENCES
Baize, H.R., & Schroeder, J.E. (1995). Personality and mate selection in personal ads: Evolutionary preferences in a public mate selection process. Journal of Social Behavior and Personality, 10, 517-536.
Betzig, L.L. (1986). Despotism and differential reproduction: A Darwinian view of history. Hawthorne, NY: Aldine.
Buss, D.M. (1988). From vigilance to violence: Tactics of mate retention. Ethology and Sociobiology, 9, 291-317.
Buss, D.M. (1989). Sex differences in human mate preferences: Evolutionary hypotheses testing in 37 cultures. Behavioral and Brain Sciences, 12, 1-49.
Buss, D.M. (1994/2003). The evolution of desire: Strategies of human mating. New York: Basic Books.
Buss, D.M. (2004). Evolutionary psychology: The new science of the mind (2nd edition). Boston: Allyn & Bacon.
Buss, D.M. (2000). The dangerous passion: Why jealousy is as necessary as love and sex. New York: Free Press.
Buss, D. M., Abbott, M., Angleitner, A., Biaggio, A., Blanco-Villasenor, A., Bruchon-Schweitzer, M [& 45 additional authors]. (1990). International preferences in selecting mates: A study of 37 societies. Journal of Cross Cultural Psychology, 21, 5-47.
Buss, D.M., & Haselton, M.G. (2005). The evolution of jealousy. Trends in Cognitive Science, 9, 506-507.
Buss, D.M., Larsen, R.J., & Westen, D. (1996). Sex differences in jealousy: Not gone, not forgotten, and not explained by alternative hypotheses. Psychological Science, 7, 373375.
Buss, D.M., Larsen, R., Westen, D., & Semmelroth, J. (1992). Sex differences in jealousy: Evolution, physiology, and psychology. Psychological Science, 3, 251-255.
Buss, D.M., & Schmitt, D.P. (1993). Sexual strategies theory: An evolutionary perspective on human mating. Psychological Review, 100, 204-232.
Buss, D.M., & Shackelford, T.K. (1997). From vigilance to violence: Mate retention tactics in married couples. Journal of Personality and Social Psychology, 72, 346-361.
Buss, D.M., Shackelford, T.K., Kirkpatrick, L.A., Choe, J., Hasegawa, M., Hasegawa, T., & Bennett, K. (1999). Jealousy and beliefs about infidelity: Tests of competing hypotheses in the United States, Korea, and Japan. Personal Relationships, 6, 125-150.
Buss, D.M., Shackelford, T.K., Choe, J., Buunk, B.P., & Dijkstra, P. (2000). Distress about mating rivals. Personal Relationships, 7, 235-243.
Clarke, R.D., & Hatfield, E. (1989). Gender differences in receptivity to sexual offers. Journal of Psychology and Human Sexuality, 2, 39-55.
Daly, M., Wilson, M., & Weghorst, S. J. (1982). Male sexual jealousy. Ethology and Sociobiology, 3, 11-27.
Darwin, C. (1859). The origin of the species. London: Murray.
Darwin, C. (1871). The descent of man and selection in relation to sex. London: Murray.
Gangestad, S.W., & Thornhill, R. (1997). Human sexual selection and developmental stability. In J.A. Simpson, & D.T. Kenrick (Eds.), Evolutionary social psychology (pp. 169-195). Mahwah, NJ: Erlbaum.
Gangestad, R.W., Thornhill, R., & Garver-Apgar, C.E. (2005). Adaptations to ovulation. In
D.M. Buss (Ed), The handbook of evolutionary psychology (pp. 344-371). New York: Wiley.
Geary, D.C., Rumsey, M., Bow-Thomas, C.C., & Hoard, M.K. (1995). Sexual jealousy as a facultative trait: Evidence from the pattern of sex differences in adults from China and the United States. Ethology and Sociobiology, 16, 355-383.
Greiling, H., & Buss, D.M. (2000). Women’s sexual strategies: The hidden dimension of extra-pair mating. Personality and Individual Differences, 28, 929-963.
Haselton, M., & Buss, D.M. (2001). The affective shift hypothesis: The functions of emotional changes following sexual intercourse. Personal Relationships, 8, 357-369.
Haselton, M., Buss, D.M., Oubaid, V., & Angleitner, A. (2005). Sex, lies, and strategic interference: The psychology of deception between the sexes. Personality and Social Psychology Bulletin, 31, 3-23.
Kenrick, D.T., Keefe, R.C., Gabrielidis, C., & Cornelius, J.S. (1996). Adolescents’ age preferences for dating partners: Support for an evolutionary model of life-history strategies. Child Development, 67, 1499-1511.
Schmitt, D.P., & Buss, D.M. (2001). Human mate poaching: Tactics and temptations for infiltrating existing relationships. Journal of Personality and Social Psychology, 80, 894-917.
Schmitt, D.P. and 118 members of the International Sexuality Description Project (2003). Universal sex differences in the desire for sexual variety: Tests from 52 nations, 6 continents, and 13 islands. Journal of Personality and Social Psychology, 85, 85-104.
Schmitt, D.P. and 121 members of the International Sexuality Description Project (2004). Patterns and universals of mate poaching across 53 nations: The effects of sex, culture, and personality on romantically attracting another person’s partner. Journal of Personality and Social Psychology, 86, 560-584.
Schmitt, D. P., Shackelford, T. K., Duntley, J., Tooke, W., & Buss, D. M. (in press). The desire for sexual variety as a tool for understanding basic human mating strategies. Personal Relationships.
Shackelford, T.K., Goetz, A., Buss, D.M., Euler, H.A., & Hoier, S. (2005). When we hurt the ones we love: Predicting violence against women from men’s mate retention. Personal Relationships, 12, 447-463.
Symons, D. (1979). The evolution of human sexuality. New York: Oxford University Press.
Tooke, W., & Camire, L. (1991). Patterns of deception in intersexual and intrasexual mating strategies. Ethology and Sociobiology, 12, 345-364.
Trivers, R.L. (1972). Parental investment and sexual selection. In B. Campbell (Ed.), Sexual
selection and the descent of man: 1871 - 1971 (pp. 136-179). Chicago: Aldine.
Wiederman, M.W., & Kendall, E. (1999). Evolution, gender, and sexual jealousy: Investigation with a sample from Sweden. Evolution and Human Behavior, 20, 121128.
Received: 02. 10. 2006.