
- Орангутаны научились использовать орудия с выгодой для себя [2019-03-22]
- Мозг собак отличил настоящие слова от тарабарщины [2019-03-22]
- Какаду Гоффина выклевали палочки из картонки и использовали их в быту [2019-03-22]
- Орангутанов уличили в создании крючков [2019-03-22]
- Любовь самок гуппи к ярким самцам объяснили генами и освещением [2019-03-22]
- Орангутаны рассказывают друг другу о прошлом [2019-03-22]
- Блеск и нищета этологии [2018-05-03]
- Почему хозяева похожи на своих собак? [2018-03-15]
- Интервью с Анатолием Протопоповым [2019-02-13]
- Цирки: развлечение vs мучение [2018-12-15]

В.С. Фридман
Врождённость (в смысле генетическую детерминированность) инстинктов не следует преувеличивать. Существенна сама способность животного создавать сложные формы видоспецифического поведения в «нужные» моменты взаимодействия, которые [формы] развиваются без обучения и вообще без какого-либо контакта с конспецификами.
При сравнении поведения разных видов «врождённость» означает типологическую определённость форм демонстраций и других комплексов фиксированных действий животного, их видоспецифичность, а не чисто генетическую детерминацию онтогенетического развития таких комплексов фиксированных действий . В анализе онтогенеза инстинктивного поведения «врождённость» означает устойчивую реализацию специфических форм соответствующего поведения в индивидуальном развитии при исчезающее малой роли социальных влияний, которые в раннем опыте животного могут «дошлифовывать» форму инстинктивного действия до видовой нормы.
. В анализе онтогенеза инстинктивного поведения «врождённость» означает устойчивую реализацию специфических форм соответствующего поведения в индивидуальном развитии при исчезающее малой роли социальных влияний, которые в раннем опыте животного могут «дошлифовывать» форму инстинктивного действия до видовой нормы.
Ещё в 1950-е гг., в дискуссии со сравнительными психологами Лоренц утверждал, что все инстинкты врождённы в первом смысле, но многие не врождённы во втором. «В теле или в поведении любого организма нет абсолютно ничего, что не зависит от внешней среды и в определенной степени не подвержено модификации благодаря окружению... То, что наследуется, - не сам признак, но пределы его модифицируемости» (Lorenz, 1956: 53)». И далее «конечно, мы не знаем, насколько то, что мы называем «врожденное поведение» прямо генетически детерминировано и насколько обязано эпигенетическим процессам... Для того, чтобы оценить модифицируемость посредством обучения, несущественно, каким образом определяется «ригидность поведения». Для этой цели нам не надо знать, как много действительно прямо зависит от генов и как много является результатом эпигенеза» (Lorenz, 1956: 67).
Например, если волчонок в подсосном периоде не будет упираться в живот матери (или вообще во что-то), и не может массировать лапками молочную железу при сосании , у него не формируются адекватные формы взрослого пищевого поведения, и в первую очередь запасание пищи. У всех этих животных заметна эмоциональная неуравновешенность, что проявлялось в сравнительно низком пороге оборонительного поведения и частых фрустрациях. Эти индивиды часто создавали в группе (с т.н. нормальными) конфликтные ситуации»
, у него не формируются адекватные формы взрослого пищевого поведения, и в первую очередь запасание пищи. У всех этих животных заметна эмоциональная неуравновешенность, что проявлялось в сравнительно низком пороге оборонительного поведения и частых фрустрациях. Эти индивиды часто создавали в группе (с т.н. нормальными) конфликтные ситуации»
Тут существенно, что для нормального формирования пищевого поведения волчат достаточно надеть на соску диск, чтобы детеныш в него упирался лапками, сделав прокол в соске таким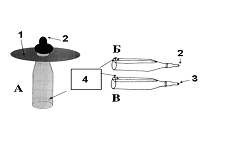 , чтобы молоко не вытекало без существенных усилий детёныша. Тот же эффект даёт использование приспособления, имитирующего тело матери
, чтобы молоко не вытекало без существенных усилий детёныша. Тот же эффект даёт использование приспособления, имитирующего тело матери » (Я.К
» (Я.К .Бадридзе. Волк. М., 2003)..
.Бадридзе. Волк. М., 2003)..
То есть в отличие от внутренних факторов (по которым наблюдается полиморфизм) социальные влияния можно смоделировать искусственно, и развившееся таким образом поведение не отличается от «естественного» по форме или по эффективности, если модель верна. Поэтому главный критерий оценки «врождённости» инстинктов – исчезающее малая роль социальных влияний и/или прошлого опыта животного на формирование соответствующих паттернов поведения после завершения онтогенеза, формирующего те же агонистические или брачные демонстрации.
В отличие от генетиков, этологов не интересует, каковая относительная роль генетических и средовых факторов в самом процессе онтогенеза, формирующем этот инстинкт как «временный орган животного» (а оно может очень сильно варьировать). То есть требования к «врождённости» поведения у этологов в норме существенно мягче «генетического»: обладал бы соответствующий паттерн поведения типологически определённой формой сразу по завершении онтогенеза, до систематических социальных влияний и какого-либо личного опыта. Всё это было чётко проговорено Лоренцем в дискуссии со сравнительными психологами по проблеме «врождённое – приобретённое в поведении» в 1950-е годы (Lorenz, 1955, 1956), но изрядно подзабылось в 1960-е, когда большинство этологов «врождённость» видоспецифического поведения стало понимать просто как «генетическую детерминированность». Это внесло изрядную путаницу, достигшую максимума в книге Р.Хайнда «Поведение животных» (см. Е.А.Гороховская «Этология. Рождение научной дисциплины», 1998).
Исследования показывают, что «полностью врождённые» формы поведения во втором смысле слова отсутствуют. Наследуется только «болванка» инстинкта – некое неопределенное действие в определённом направлении (в сторону партнёра – потенциального источника ключевых раздражителей, запускающих инстинктивный акт). Далее идут «шлифовка» и «оттачивание» инстинкта в актах употребления во взаимодействии с «нужным» социальным компаньоном, в процессе которой движения обретают видоспецифическую форму, а животное всё точней ориентируется по «пусковым» сигналам извне, всё лучше выделяет сигналы из многообразия окружения и т.д.
Даже у низших позвоночных естественное окружение животного - такое же обязательное условие развития инстинкта, как его генетически детерминированная основа. У высших позвоночных, у птиц и млекопитающих развитие инстинктивных форм поведения (в первом смысле) прямо требует социальных влияний для «доводки» соответствующего поведения до дефинитивной формы и работоспособного состояния.
Без опыта употребления инстинкта в специфической социальной ситуации, накопления истории «проб и ошибок» запуска инстинктивного акта «созревание» дефинитивной формы последнего или не происходит вовсе (птицы и млекопитающие) или «созревший» употребляется слишком неспецифически и неточно, чтобы быть эффективным. Продуктивная аналогия: сходным образом профермент трансформируется в активный фермент другими ферментативными системами организма.
Это касается даже таких простейших реакций, как клевание цветного пятнышка на конце клюва родителя, которое только что обсохший чайчонок совершает, чтобы побудить взрослых птиц отрыгнуть кормовой комок. Затем его птенец склёвывает, то есть одно и то же действие (движение) выбора ориентира, прицеливания и клевка поочередно используется в специализированном варианте социальной активности (реакция на цветное пятно как на сигнал, устанавливающий взаимодействие птенца с компаньоном-родителем), и затем в чисто функциональном (потребление пищи сами птенцом).
Эта реакция впервые описана Н.Тинбергеном и А.Пердеком у серебристой чайки Larus argentatus (Tinbergen, Perdeck, 1950), и вошла во все учебники этологии как «парадный пример» инстинктивного акта, в том числе в отношении врождённости действия (Tinbergen, 1951; Хайнд, 1975). Её переисследование на примере аналогичной реакции клевания смеющейся чайки  Larus atricilla
Larus atricilla Дж.Хэйлменом показало более чем существенную «доводку» формы движения, его ориентированности на внешний сигнал (и, следовательно, точности и эффективности) за счёт социальных влияний и опыта употребления реакции в прошлом (Hailman, 1967, 1968, 1971).
Дж.Хэйлменом показало более чем существенную «доводку» формы движения, его ориентированности на внешний сигнал (и, следовательно, точности и эффективности) за счёт социальных влияний и опыта употребления реакции в прошлом (Hailman, 1967, 1968, 1971).
Во-первых, действительно ли у птенцов до начала попыток клевания уже сформирован образ ключевого стимула, то есть они различают голову родителя, клюв, маленькое цветное пятнышко на нём, и предпочитают клевать именно пятно, а не другие части клюва, или пятно на клюве предпочитают пятну, скажем, на лбу модели? Оказывается, нет, этот образ формируется при взаимодействии птенцов с родителями и друг с другом в первые дни жизни.
В классических экспериментах Тинбергена и Пердека модель держали в руке, поэтому при ее маятникообразном движении вместе с кистью руки пятно на лбу двигалось медленнее и по более короткой дуге, чем на клюве. Кроме того, птенцу приходилось тянуться вверх, чтобы клюнуть пятно на лбу. Поэтому чайчата пятно на клюве клевали чаще и точней, чем пятно на лбу, что интерпретировалось как априорное различение стимула.
Хэйлмен повторил эти опыты, но добавил еще одну модель с цветным пятном на лбу, закреплённую на стержне таким образом, чтобы лобное пятнышко было на том же расстоянии от оси вращения, как пятно на клюве другой модели. Экспериментальная установка также имела подвижный пол, высоту которого изменяли так, чтобы красное пятно на модели оставалось всегда на уровне глаз птенца. Эту модель назвали «быстрой», так как пятно у нее на лбу перемещалось быстрее, чем в «медленной» модели Тинбергена и Пердека.
Результаты неутешительны для защитников идеи о безусловной врожденности инстинктов: для только что вылупившихся птенцов «быстрая» модель с пятном на лбу столь же эффективна, как «медленная» с пятном на клюве. Затем чайчат вернули в гнездо и повторно тестировали на 3-й и 7-й дни жизни (в природе клевание пятна полностью формируется на 7-8 день). Птенцы существенно улучшили результат и определённо предпочитали пятно на клюве (Hailman, 1967).
Существенно улучшается также точность клевков. Для этого исследования птенцам предъявляли схематическое изображение головы взрослой чайки на маленьких карточках. Будучи прикреплёнными к подвижному стержню, модели перемещались в горизонтальном направлении перед глазами птенца, а он их клевал.
Яйца чаек инкубировали в полной темноте, и до начала опытов птенцы не получали никаких зрительных стимулов. Сразу после вылупления каждому птенцу позволяли 10-15 раз клюнуть подвижную двумерную модель головы взрослой птицы, обозначая точками места клевков. Протестированных птенцов метили и возвращали в гнездо (то есть он естественным образом воспитывался приёмными родителями) и брали взамен яйцо, близкое к вылуплению. На 1-й, 3-й и 5-й дни жизни половину меченых птенцов тестировали., другую половину – на 2-й, 4-й и 6-й день, и после опытов сразу возвращали в гнездо. Сразу после вылупления только треть их клевков достигает цели. Но затем результат быстро улучшается: на следующий день точность >50%, через два дня после вылупления - > 75% и дальне не меняется. Разброс данных, особенно горизонтальный, у «среднего» птенца очень быстро падает с возрастом (Hailman, 1967, 1968).
Какой механизм ответственен за повышение точности клевков? Для ответа на вопрос разработали более сложный эксперимент с двумя контрольными группами птенцов, выращенных в естественных условиях. Чтобы птенцы экспериментальных групп не приобрели опыта зрительной координации клевания, их держали в темных брудерах. Одну группу кормили искусственно, вторая два дня вообще не получала пищи и жила за счет обильных жировых запасов. Чтобы выяснить, влияет ли на точность клевков активность, связанная с вылуплением, птенцов третьей группы сразу после появления эмбриональных писков извлекали из яйца и помещали в инкубатор.
Всем птенцам предъявляли модель головы взрослой птицы, реакции чайчат регистрировали на кинопленке, что позволило установить % точных клевков. Повышение точности отмечено во всех пяти группах, но лишь в двух контрольных оно достигает нормы (>75%). Если зрительная депривация существенно тормозит рост точности клевания, исключение активности, связанной с вылуплением, практически не значимо. Следовательно, зрительный опыт необходим для максимизации точности клевков, но некоторое улучшение происходит и при зрительной депривации, возможно за счёт улучшения позных реакций (Hailman, 1968).
Далее исследовали, как птенец начинает выбирать оптимальное расстояние для точного клевка (с учётом собственного роста и реакций взрослой особи). Здесь действует механизм саморегуляции, основанный на восприятии расстояния. Если неопытный птенец встанет слишком близко к модели, то удар отбросит его назад на 2-3 см. Если он будет слишком далеко, он не достает до нее при клевке и падает вперед на 4-5 см. Столь серьёзные ошибки старшие птенцы допускают очень редко: оптимальное расстояние устанавливается во взаимодействиях со взрослыми и с другими птенцами (см.ниже).
Во-вторых, как птенец научается поворачивать голову так, чтобы точно схватить клюв взрослой птицы, и нацеливается именно на кончик клюва, чтобы получить пищу? С одной стороны, в основе голода лежит нечто врожденное. У сытых птенцов частота клевков возрастает с увеличением интервала времени после кормления. С другой, та же картина отмечена у птенцов, вылупившихся в темноте, и искусственно накопленных досыта на второй день жизни (после чего их тестировали также как первых).
Детали движений птенца при клевании анализировали с помощью высокоскоростной киносъемки. Полностью развитая пищевая реакция птенца в возрасте нескольких дней включает четыре компонента: 1) открывание и последующее закрывание клюва, 2) движение головы вверх и вперед по направлению к голове взрослой птицы, затем вниз и назад, 3) поворот головы в сторону в расчете на то, чтобы захватить клювом клюв взрослой птицы, и обратный поворот головы, 4) легкий толчок ногами вверх и вперед.
Покадровый анализ фильма выявил сильную изменчивость временных отношений между этими компонентами, как у одних и тех же птенцов в разных актах клевания, так и между птенцами. С возрастом эти вариации слегка уменьшаются (возможно, из-за улучшения координации движений), но не сильно, а вот точность попадания в цветное пятнышко или схватывания клюва матери сильно растёт.
У птенцов, выросших в естественных условиях, поворот головы в сторону с возрастом происходит все чаще. Чтобы выяснить, насколько это зависит от зрительного опыта, анализировали фильмы, снятые во время тестов на точность клевания, по всем пяти группам птенцов. Оказалось, что у птенцов, не имевших опыта клевания, сколько-нибудь заметный поворот головы отмечается очень. У птенцов в природе в первый день жизни поворотов головы почти бывает, но затем они появляются быстро совершенствуются.
При неточных клевках птенца с раскрытым клювом, клюв взрослого, расположенный не строго вертикально, часто оказывается между челюстями птенца. Тогда движение последнего по инерции вперед поневоле приводит к повороту в сторону. Возможно, именно так птенец научается поворачивать голову, чтобы схватить клюв взрослой птицы (Hailman, 1968, 1971).
И самое главное – как совершенствуется распознавание образов, запускающих реакцию клевания? Ведь следует точно распознавать родителей, на родителе клюв, на клюве – цветное пятно, и клевать туда, а не в лоб и не в лапы (у смеющейся чайки они красные также как и клюв). Развитие этой реакции идёт по знакомой схеме: в природе птенцы иногда клюют не только клюв взрослой птицы, но и иные предметы, в том числе другие части тела родителя. Правда, у более старших птенцов всё больший процент клевков ориентируется на клюв и на кончик клюва.
То есть недавно вылупившиеся птенцы представляют себе родителей довольно смутно, но с возрастом, по мере накопления опыта, образ взрослой птицы делается все более четким. Эти процессы исследовали с помощью серии «разборных» моделей головы, у которых изменяли или устраняли отдельные части, чтобы выявить наиболее эффективные стимулы пищевой реакции (Hailman, 1967, 1971).
Модель закрепляли на стержне, колеблющемся в такт с метрономом, так что скорость движения модели была известна. В каждом опыте птенцу предъявляли пять моделей в случайном порядке, и регистрировали все клевки в последующие 30 с. Требовалось выявить наиболее эффективные стимулы для птенцов, до начала эксперимента находившихся в темноте первые сутки после вылупления.
В первых трех опытах птенцы одинаково реагировали на все модели, кроме той, у которой не было клюва. Следовательно, новорожденные птенцы реагируют в основном на клюв родителей, а не на особенности головы (даже само её наличие). Какой именно клюв предпочитается, исследовали с помощью деревянных палочек, окрашенных в красный цвет. Палочки предъявляли в горизонтальном или вертикальном положении: в обоих случаях она либо была неподвижна, либо перемещалась в горизонтальном / вертикальном направлении. Всякая вертикальная палочка вызывала клевание чаще, чем любая горизонтальная, из вертикальных наиболее эффективна модель, движущаяся в горизонтальном направлении.
Это хорошо согласуется с естественным окружением птенца чайки в гнезде, когда вертикально расположенный клюв взрослой птицы часто движется горизонтально перед его глазами. Именно этот образ и распознаётся как пусковой для реакции клевания, в первые дни несовершенной и неточной и, соответственно, врождённым является именно он. Дальнейшее совершенствование реакции – результат именно социальных влияний (Hailman, 1967, 1968, 1971).
Далее оказалось, что вертикально движущийся вертикальный стимул был не эффективнее неподвижного вертикального. И горизонтальное, и вертикальное перемещение горизонтального стимула было одинаково действенно и, как это ни странно, превосходило по эффективности неподвижный горизонтальный стимул. Отсюда следует, что адекватными стимулами пищевой реакции птенцов в равной мере служат оба типа движения - горизонтальное перемещение и движение в направлении, перпендикулярном продольной оси стимула.
Это позволяет объяснить, почему неподвижная вертикальная палочка столь же неэффективна, что и вертикальная палочка, движущаяся вертикально (в обоих случаях нет движений указанных типов). В случае горизонтальной палочки вертикальное движение перпендикулярно продольной оси стимула и потому так же эффективно, как горизонтальное, и реакция здесь при обоих движениях более интенсивна, чем при полной неподвижности стимула (Hailman, 1967).
На следующем этапе испытывали вертикальные стимулы разных диаметров, движущиеся с пятью разными скоростями. Наиболее эффективна была палочка 8 мм в диаметре независимо от скорости движения. Соответственно, наиболее эффективна скорость около скорость 12 см/с независимо от диаметра палочки.
Затем тестировали семидневных птенцов. В отличие от птенцов в первые дни жизни, они оказались предельно чувствительны даже к незначительным изменениям формы головы и клюва. Они также легко модели головы птиц своего вида от моделей серебристых чаек, и клюют только модели достаточно сходные с взрослыми конспецификами.
.Это показывает, насколько точно «созревание» пищевой реакции птенцов в нормальном окружении подгоняется к размерам стимулов. Толщина клюва взрослой чайки в вертикальном направлении 10,6 мм, в горизонтальном - 3,1 мм, так что средняя толщина как раз около 8 мм, средняя скорость движения клюва при кормлении птенцов в природе около 14,5 см/с. Вертикальная палочка, находящаяся выше уровня глаз птенца, гораздо эффективнее, чем такой же стимул, предъявленный внизу. Также предпочтение отдается наклонным объектам, что снижает вероятность реакции на ноги родителей.
При изменении данных параметров, скажем, в ходе эволюционного процесса развитие пищевой реакции птенцов теми же механизмами легко подгоняется к другим размерам клюва и другой экспрессии движений родителей. Поэтому «полностью врождённые инстинктивные акты просто вредны, как всякая избыточная ригидность организации, а действительно врождённые акты должны быть предельно просты и достаточно неопределённы в совершении действия и в выборе пускового стимула, иначе не допускают «подгонки» механизмами, исследованными J.P.Hailman (1967, 1968, 1971).
Механизмы улучшения всех реакций птенца связан не с классическим обучением, а с обострением восприятия, вызванным солидарными действиями компаньонов – родителей и других птенцов.
В тех же опытах исследовали, узнают ли только что вылупившиеся птенцы пищу, когда сталкиваются с нею? Чтобы выяснить это, поставили четыре чашечки с кормом в углах небольшого ящика и пронаблюдали, как выведенные в инкубаторе птенцы первый раз в жизни находят пищу. Время, затраченное птенцом на поиски пищи, оказалось обратно пропорциональным частоте клевков. Этого и следовало ожидать, если исходить из того, что птенцы разыскивают пищу только методом проб и ошибок.
Но если им дать насытиться и пересадить в другое место, пока они не проголодаются, то в повторном опыте они находят пищу гораздо быстрее. В третьем необходимое время сокращается до минимума. Это нельзя объяснить увеличением частоты клевков, так как во втором в последующих опытах она лишь незначительно выше, чем в первом.
Следовательно, птенцы очень быстро научаются распознавать пищу или место, где её стоит брать. Один из механизмов этого - птенец не клюет пищу, то взрослая птица обычно берет ее в клюв. При первом кормлении птенец продолжает клевать клюв родителя и после того, как срыгнутая пища падает на дно гнезда. В конце концов, взрослые подбирают пищу, и птенец натыкается на нее при клевке, нацеленном на клюв родителя.
Это говорит об адаптивности вылупления птенцов именно с «несовершенной» реакцией клевания – промахи из-за малой точности клевков в первые дни жизни повышают вероятность того, что птенец случайно натолкнется на пищу.
В том же направлении «работает» и взаимодействие между птенцами. Клевки одного чайчонка часто нацелены на белый кончик клюва другого (опять же из-за несформированности системы распознания образов, необходимых для «запуска» инстинктивного акта). Птенцы вылупляются с интервалом около 12 часов, и к моменту вылупления следующего старший уже получает корм.
Во время такой кормёжки младший обычно пытается клевать клюв старшего, и тоже натыкается на пищу, и одновременно «шлифует» само движение, подгоняя его по более точному образцу у собрата. Наблюдения в природе показывают, что птенцы Larus atricilla первый раз находят корм именно так.
Безусловно, это совершенствование инстинкта под воздействием специфических социальных влияний. Однако это не обучение (формирование навыка), а именно созревание инстинкта, так как «улучшение навыков» демонстрирования в следующем моменте процесса развития поведения не связано с интенсивностью (или успешностью) «упражнения» в прошлом (тут я не согласен с J.P.Hailman). При настоящем обучении подобная связь всегда имеется и сильна (Мак-Фарленд, 1988; Резникова, 2005), а сформированный навык поддаётся забыванию и переделке (Резникова, 2003). Далее, в процессе «созревания» инстинкта нет прямо пропорциональной зависимости между долей правильного подкрепления в прошлые моменты времени и уменьшением ошибок в следующий период, она носит пороговый характер и пр. (Hailman, 1967, 1971).
Источники:
Резникова Ж.И., 2005. Интеллект и язык животных и человека. Основы когнитивной этологии. М.: Академкнига. 518 с.
Резникова Ж. И., 2006. Исследование орудийной деятельности как путь к интегральной оценке когнитивных возможностей животных// Журнал общей биологии. Т.67. № 1. С.2-22
Hailman J. P. 1967. The ontogeny of an instinct: the pecking response in chicks of the laughing gull (Larus atricilla L.) and related species // Behav. Suppl. Vol. XV. E.J. Brill, Leiden. 196 pp.
Hailman J. P., 1968. Visual-cliff responses of newly-hatched chicks of the laughing gull Larus atricilla// Ibis. Vol.101. P.187-200.
Hailman J. P. 1971. The role of stimulus-orientation in eliciting the begging response from newly-hatched chicks of the laughing gull (Larus atricilla)// Anim. Behav. Vol.19. P.328-335.
Lorenz K., 1955. Morphology and behavior patterns in closely allied species // Transactions of the First Conference on Group Processes (1954) / Ed. by B. Schaffner. N.Y.: Macy Foudation. P. 168-220.
Lorenz K, 1956. The objectivistic theory of instinct // L'instinct dans le comportement des animaux et de l'homme / Dir. par rasse, P. P. Paris: Masson et Cie. P. 51-76.